
Page 53 - 《广西植物》2020年第8期
P. 53
1 1 0 8 广 西 植 物 40 卷
竞争所致ꎻ李娟等(2019) 认为植物从幼龄阶段到
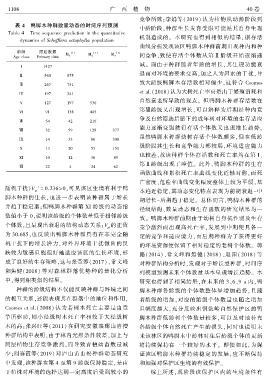
表 4 鸭脚木种群数量动态的时间序列预测
中龄阶段ꎬ种群生长发育受限可能是其自身生理
Table 4 Time sequence prediction in the quantitative
机制造成的ꎮ 本研究也得到相似的结果ꎬ据存活
dynamics of Schefflera octophylla population
曲线分析发现该区鸭脚木种群前期出现种内和种
龄级 原始数据 [1] [1] [1]
M 3 M 6 M 9 间竞争ꎬ致使存活个体数从第Ⅱ龄级开始逐渐递
Age class Primary data
减ꎮ 而由于种群随着年龄的增长ꎬ其生理功能衰
I 1427
退而对环境的要求变高ꎬ加之人为因素的干扰ꎬ导
Ⅱ 560 875
致大龄级鸭脚木存活数相对偏少ꎬ这符合 Coomes
Ⅲ 267 751
et al.(2018)认为大树死亡率高是由于繁殖消耗和
IV 197 341
自然衰老所导致的观点ꎮ 但鸭脚木种群存活数在
V 127 197 536
第Ⅻ龄级又出现增长ꎬ可以解释为后期经种内竞
VI 91 138 445
争及自然筛选后留下的成年树对环境的生存适应
Ⅶ 54 42 216
能力逐渐变强使得存活个体数又出现增长趋势ꎮ
Ⅷ 32 59 128 377
虽然鸭脚木种群幼树存活个体数颇多ꎬ但在低龄
IX 14 33 86 308
级阶段其生长和竞争能力都较弱ꎬ环境适应能力
X 13 20 55 151
也较差ꎬ故该种群个体存活数和死亡率均在第 I、
XI 10 12 36 89
第Ⅱ龄级出现了峰值ꎮ 此外ꎬ鸭脚木种群的生存
Ⅻ 22 4 24 62
函数曲线和累积死亡率曲线变化近轴对称ꎬ而死
亡密度、危险率曲线变化幅度整体上较为平缓ꎬ基
随机干扰)V ′ = 0.236>0ꎬ可见该区生境有利于鸭 本趋近稳定ꎬ其动态变化特点表现为前期衰退-中
pi
脚木种群的生长ꎬ也进一步表明该种群属于增长
期增长-后期趋于稳定ꎮ 总体而言ꎬ鸭脚木种群的
并趋于稳定型ꎻ但鸭脚木种群第 XI 龄级的动态指
径级结构、数量动态和生存函数的研究结果均一
数值小于 0ꎬ说明该龄级的个体数量低于相邻龄级
致ꎮ 鸭脚木种群前期由于幼树自身抗性弱及生存
个体数ꎬ且呈现出衰退的结构动态关系ꎻV 的正值 竞争激烈而出现高死亡率ꎬ发展到中期便具备一
pi
为 30.685ꎬ也反映出鸭脚木种群具有在非完全随 定的竞争和适应能力ꎬ而后期种群为了获得更好
机干扰下的增长潜力ꎬ对外界环境干扰做出的反 的环境资源便保留了相对稳定的老树个体数ꎮ 韩
映较为敏感且能很好地适应该区的生长环境ꎬ形 路(2014)、常文峰和焦健(2018)、赵阳(2018) 等
成了良好的生存策略ꎬ这与张萱蓉(2017)、常文峰 对种群结构分析时ꎬ发现对于增长型种群ꎬ时间序
和焦健(2018) 等对森林群落优势种的量化分析 列模型预测未来个体数量基本呈现增长趋势ꎮ 本
中ꎬ得到相类似的结果ꎮ 研究也得到了相同结果ꎬ在未来的 3、6、9 a 内ꎬ鸭
种群的龄级结构不仅能反映种群与环境之间 脚木种群各龄级的个体数整体呈增加趋势ꎬ且随
的相互关系ꎬ还能表现其在群落中的地位和作用ꎬ 着龄级的增加ꎬ对应的龄级个体数量也随之增加
Coomes et al.(2008) 认为若树木死亡主要是由竞 且幅度越大ꎬ充分反映出铜鼓岭自然保护区的鸭
争所驱动ꎬ则小径级树木死亡率相较于大径级树 脚木种群低龄树个体数量较多ꎬ可以及时地补充
木的高ꎻ张兴旺等(2011) 在研究安徽琅琊山青檀 各龄级个体自然死亡产生的损失ꎬ同时也说明未
种群结构中表明ꎬ由于林内光照条件较差ꎬ加上与 来该林区的鸭脚木中龄林对往后龄级个体的更新
同层植物生存竞争激烈ꎬ而导致青檀幼苗数量减 将持续保持在一个较好的水平ꎮ 即便如此ꎬ为保
少ꎻ周赛霞等(2019) 对庐山黄山松种群动态研究 证该区鸭脚木种群持续稳定的发展ꎬ应不断保持
中发现ꎬ该种群在第 4 至第 9 龄级保持稳定ꎬ是由 和加强对保护区生境的有效保护ꎮ
于植株对环境的选择达到一定高度后受到较小的 综上所述ꎬ现阶段该保护区内的生境条件有