
Page 101 - 《广西植物》2022年第3期
P. 101
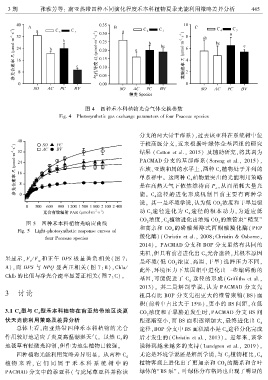
3 期 张雅芳等: 南亚热带四种不同演化程度禾本科植物夏季光能利用策略差异分析 4 4 5
图 4 四种禾本科植物光合气体交换参数
Fig. 4 Photosynthetic gas exchange parameters of four Poaceae species
分支的两大骨干群系)ꎬ过去该亚科在系统树中位
于较高级分支ꎬ近来根据叶绿体全基因组的研究
结果 (Cotton et al.ꎬ 2015) 及辅助研究ꎬ将其调为
PACMAD 分支的基部群系( Soreng et al.ꎬ 2015)ꎬ
在族、亚族和属的水平上ꎬ两种 C 植物处于并列的
4
单系群中ꎮ 这两种 C 植物最突出的光能利用策略
4
是在高热天气下依然维持高 P ꎬ从而消耗大量光
n
能ꎮ C 途径的进化形成机制目前主要有两种学
4
说ꎮ 其一是环境学说ꎬ认为低 CO 浓度和干旱是驱
2
动 C 途径进化为 C 途径的根本动力ꎬ为适应低
3 4
CO 浓度ꎬC 植物进化出浓缩 CO 的维管束“ 鞘泵”
图 5 四种禾本科植物光响应曲线 2 4 2
和高亲和 CO 的磷酸烯醇式丙酮酸羧化酶( PEP
Fig. 5 Light ̄photosynthetic response curves of 2
four Poaceae species 羧化酶)(Christin et al.ꎬ 2008ꎻChristin & Osborneꎬ
2014)ꎮ PACMAD 分支和 BOP 分支虽然有共同的
先祖ꎬ但只有前者进化出 C 光合途径ꎬ其根本原因
4
果显示ꎬF / F 和正午 DPS 极显著负相关( 图 7:
v
m
是环境( 低 CO 浓度、高温、干旱) 选择压力不同ꎬ
2
A)ꎬ而 DPS 与 NPQ 显著正相关( 图 7:B)ꎬChla /
此外ꎬ环境压力下基因组中进化出一些编码酶的
Chlb 的比值与净光合速率显著正相关(图 7:C)ꎮ
基因ꎬ可能促进了 C 途径的形成( Griffiths et al.ꎬ
4
2013)ꎮ 其二是解剖学说ꎬ认为 PACMAD 分支先
3 讨论 祖具有比 BOP 分支先祖更大的维管束鞘( BS) 面
积(前者中占比大于 15%)ꎬ更小的 BS 间距ꎬ在低
3.1 C 型与 C 型禾本科植物在南亚热带地区炎夏 CO 浓度和干旱胁迫发生时ꎬPACMAD 分支 BS 间
4
3
2
伏天光能利用策略及差异分析 距逐渐变小ꎬ而 BS 面积逐渐加大ꎬ最终进化出 C
4
总体上看ꎬ南亚热带四种禾本科植物的光合 途径ꎬBOP 分支中 BS 面积减小是 C 途径分化完成
4
作用较好地适应了炎夏高温潮湿天气ꎮ 虽然 C 的 后才发生的( Christin et al.ꎬ 2013)ꎮ 近年来ꎬ该学
4
地毯草有轻微光抑制ꎬ但作为地生植物已较强ꎮ 说得到越来越多的支持( Lundgren et al.ꎬ 2019)ꎮ
四种植物光能利用策略差异明显ꎮ 从两种 C 无论是环境学说还是解剖学说ꎬ与 C 植物相比ꎬC
4 3 4
植 物 来 看ꎬ 它 们 同 属 于 禾 本 科 系 统 树 中 的 植物客观上进化出了更加亲和 CO 的酶系和含叶
2
PACMAD 分支中的黍亚科( 与虎尾草亚科并称该 绿体的“BS 泵”ꎬ 叶绿体分布格局也出现了明显的