
Page 103 - 《广西植物》2022年第3期
P. 103
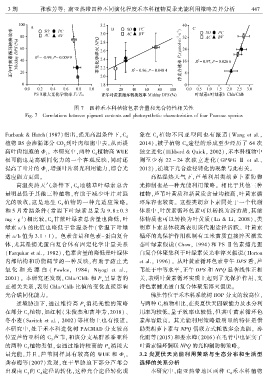
3 期 张雅芳等: 南亚热带四种不同演化程度禾本科植物夏季光能利用策略差异分析 4 4 7
图 7 四种禾本科植物色素含量和光合特性相关性
Fig. 7 Correlations between pigment contents and photosynthetic characteristics of four Poaceae species
Furbank & Hatch(1987)指出ꎬ强光高温条件下ꎬC 象在 C 植物不同亚型间也有报道( Wang et al.ꎬ
4 4
植物 BS 会渗漏部分 CO 到叶肉细胞中去ꎬ从而提 2014)ꎬ被子植物 C 途径的形成至少经历了 64 次
2 4
高叶肉细胞的 Φ ꎮ 本研究中ꎬ两种 C 植物高 WUE 独立进化(Hibberd & Quickꎬ 2002)ꎬ禾本科植物中
i 4
极可能也是高碳同化力的一个客观反映ꎬ同时还 则至 少 有 22 ~ 24 次 独 立 进 化 ( GPWG Ⅱ et al.ꎬ
提高了叶片的 Φ ꎬ增强叶片弱光利用能力ꎬ综合光 2012)ꎬ逆境下光合途径转化的现象与此有关ꎮ
i
适应能力更强ꎮ 高温湿热天气下ꎬ芦苇利用类胡萝卜素防御
高温炎热天气条件下ꎬC 地毯草叶绿素总含 光抑制也是一种光能利用策略ꎮ 相比于其他三种
4
量明显低于其他三种植物ꎬ有助于减少叶片对强 植物ꎬ芦苇叶黄质和新黄质含量均较高ꎬ叶黄素循
光的吸收ꎬ这是地生 C 植物的一种光适应策略ꎮ 环库容也较高ꎮ 这些类胡萝卜素同处于一个代谢
4
和 5 月常温条件( 常温下叶绿素总量为 9.8 ± 0.3 体系中ꎬ叶黄素循环色素可以转换为新黄质ꎬ其前
mgg )相比较ꎬC 甘蔗叶绿素总含量也降低ꎬ叶 体物质也可以转换为叶黄质( Lu & Liꎬ 2008)ꎬ类
 ̄1
4
绿素 a / b 的比值也略低于常温条件( 常温下叶绿 胡萝卜素总体较高表明该代谢途径活跃ꎮ 叶黄素
素 a / b 值为 3.1 ∶ 1)ꎮ 色素含量和色素-蛋白复合 循环的光保护作用机制有玉米黄质直接淬灭激发
体ꎬ尤其是捕光蛋白复合体有固定化学计量关系 态叶绿素假说( Chowꎬ 1994) 和 PS Ⅱ色素捕光蛋
(Farquhar et al.ꎬ 1982)ꎬ色素含量的降低是叶绿体 白复合体聚集利于叶绿素荧光非淬灭假说( Horton
内部结构和功能调节的一种反映ꎬ有助于防止光 et al.ꎬ 1996)ꎮ 从叶黄素循环色素中午 DPS 看ꎬ芦
氧 化 和 光 漂 白 ( Powlesꎬ 1984ꎻ Niyogi et al.ꎬ 苇处于中等水平ꎬ正午 DPS 和 NPQ 显著线性正相
2001)ꎮ 本研究还发现ꎬChla / Chlb 和 P 呈显著的 关ꎬ表明叶黄素循环实质上起到了光保护作用ꎬ支
n
正相关关系ꎬ表明 Chla / Chlb 比值的变化直接影响 持色素捕光蛋白复合体聚集淬灭假说ꎮ
光合碳同化能力ꎮ 佛肚竹位于禾本科系统树 BOP 分支的较高位ꎬ
逆境胁迫下ꎬ通过维持高 P 消耗光能的策略 与两种 C 植物相比ꎬ在炎夏伏天固碳能力及水分利
n
4
在部分 C 植物ꎬ如红树( 朱俊杰和曹坤芳ꎬ2018)、 用率均较低ꎬ量子效率也较低ꎬ但其叶黄素循环色
3
冬小麦( Savitch et al.ꎬ 2002) 等植物上也有报道ꎮ 素库容最高ꎮ 其光能利用策略最明显的特征是借
本研究中ꎬ处于禾本科进化树 PACMAD 分支较高 助类胡萝卜素与 NPQ 偶联方式耗散多余光能ꎮ 孙
位置芦竹亚科的 C 芦苇ꎬ和该分支基群系黍亚科 化雨等(2015) 和娄永峰(2016) 在毛竹中也证实了
3
的两种 C 植物类似ꎬ也通过维持较高的 P 消耗大 叶黄素循环偶联 NPQ 的光抑制防御策略ꎮ
4 n
量光能ꎬ并且ꎬ芦苇同样具有较高的 WUE 和 Φ ꎮ 3.2 炎夏伏天光能利用策略与生态分布和生活型
i
龚春梅等(2007)发现ꎬ在干旱胁迫下部分芦苇会 选择的关系分析
出现由 C 向 C 途径的转化ꎬ这种光合途径转化现 本研究中ꎬ南亚热带地区两种 C 禾本科植物
3 4 4