
Page 127 - 《广西植物》2025年第11期
P. 127
11 期 胡晓玉等: 转录组分析马缨杜鹃响应高温胁迫的分子机制 2 0 7 1
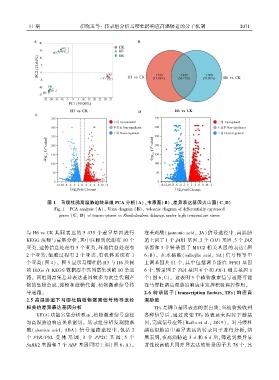
图 1 马缨杜鹃高温胁迫转录组 PCA 分析(A)、韦恩图(B)、差异表达基因火山图(C、D)
Fig. 1 PCA analysis (A)ꎬ Venn diagram (B)ꎬ volcanic diagram of differentially expressed
genes (Cꎬ D) of transcriptome in Rhododendron delavayi under high temperature stress
与 H6 vs CK 共同表达的 5 435 个差异基因进行 在茉莉酸(jasmonic acidꎬ JA) 信号途径中ꎬ高温胁
KEGG 注释与富集分析ꎬ其中注释到代谢有 10 个 迫上调了 1 个 JAR1 基因、2 个 COI1 基因、5 个 JAZ
亚类ꎬ遗传信息处理有 5 个亚类ꎬ环境信息处理有 基因和 3 个转录因子 MYC2 相关基因的表达( 图
2 个亚类ꎬ细胞过程有 2 个亚类ꎬ有机体系统有 1 6:B)ꎮ 在水杨酸( salicylic acidꎬ SA) 信号转导中
个亚类(图 4)ꎮ 图 5 显示马缨杜鹃 H3 与 H6 共同 上调基因共 11 个ꎬ其中包括调节蛋白 NPR1 基因
的 DEGs 在 KEGG 数据库中共同富集到的 10 条通 6 个、转录因子 TGA 基因 4 个和 PR ̄1 相关基因 1
路ꎮ 两组间富集差异表达基因较多为次生代谢产 个(图 6:C)ꎮ 这表明 3 个植物激素信号通路可能
物的生物合成、淀粉和蔗糖代谢、植物激素信号转 在马缨杜鹃高温胁迫响应中发挥积极调控作用ꎮ
导通路ꎮ 2.6 转录因子( transcription factorsꎬTFs) 响应高
2.5 高温胁迫下马缨杜鹃植物激素信号转导途径 温胁迫
相关的差异表达基因分析 TFs 是调节基因表达的蛋白质ꎬ当植物接收到
KEGG 功能富集分析显示ꎬ植物激素信号途径 各种信号后ꎬ通过改变 TFs 的表达来调控下游基
与高温胁迫响应关系密切ꎮ 转录组分析发现脱落 因ꎬ完成信号应答(Baillo et al.ꎬ 2019)ꎮ 对马缨杜
酸(abscisic acidꎬ ABA) 信号通路途径中ꎬ包括 2 鹃高温胁迫中差异表达的转录因子进行分析ꎬ结
个 PYR / PYL 受 体 基 因、 3 个 PP2C 基 因、 5 个 果表明ꎬ在高温胁迫 3 d 和 6 d 后ꎬ筛选到差异显
SnRK2 基因和 7 个 ABF 基因同时上调( 图 6:A)ꎮ 著性较高的共同差异表达的转录因子共 78 个ꎬ其