
Page 138 - 《广西植物》2025年第12期
P. 138
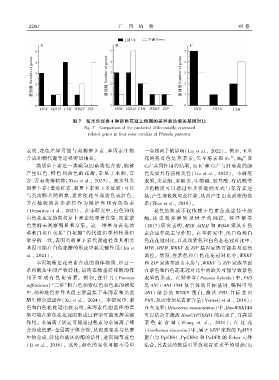
2 2 8 0 广 西 植 物 45 卷
图 7 帕米尔报春 4 种颜色花冠上预测的差异表达相关基因对比
Fig. 7 Comparison of the predicted differentially expressed
related genes in four color corollas of Primula pamirica
表明ꎬ花色差异可能与类胡萝卜素、苯丙素生物 一金属离子的影响(Liu et al.ꎬ 2022)ꎮ 例如ꎬ玉米
2+
3+
合成和糖代谢等途径密切相关ꎮ 花的亮 蓝 色 是 芹 菜 素、矢 车 菊 素 和 Fe 、 Mg 及
类胡萝卜素是一类碳氢四萜类化合物ꎬ能够 Ca 共同作用的结果ꎬ而 K 和 Cu 与红菊花的颜
2+
+
2+
产生红 色、 橙 色 和 黄 色 的 花 瓣ꎬ 常 见 于 水 仙、 百 色发展具有强相关性( Liu et al.ꎬ 2022)ꎮ 本研究
合、万寿菊等植物( Xiao et al.ꎬ 2023) ꎮ 疏水性类 表明ꎬ类黄酮、多酚类、生物碱、氨基酸、有机酸等
胡萝卜素( 番茄红素、胡萝卜素和玉米黄质) 可以 共色物质可以通过非共价键的方式与花青素连
与类黄酮共同积累ꎬ通常使花呈现黄色或红色ꎬ 接ꎬ产生增色效应及红移ꎬ从而产生由蓝到紫的色
并在植 物 的 许 多 部 位 作 为 保 护 性 和 着 色 色 素 系(Zhao et al.ꎬ 2016)ꎮ
( Demurtas et al.ꎬ 2023) ꎮ 在本研究中ꎬ白色和浅 花色的形成不仅依赖于 色 素 合 成 途 径 中 的
白色花花冠的类胡萝卜素途径显著富集ꎬ而在紫 酶ꎬ 还 受 到 多 种 转 录 因 子 的 调 控ꎮ 侯 泽 豪 等
色类群未观察到显著富集ꎮ 这一结果与黄花油 (2017) 研究表明ꎬMYB、bHLH 和 WD40 家族在色
菜和白花自交系“ 白花瓣” 的代谢组学和转录组 素合成中起主导作用ꎮ 在本研究中ꎬ浅白色和白
学分析一致ꎬ表明类胡萝卜素代谢途径及其相关 色花花冠对比ꎬ以及浅紫色和白色花花冠对比中ꎬ
基因可能在白色花瓣的形成中起关键作用( Jia et MYB、bHLH、WRKY 及 ZIP 基因家族可能参与花色
al.ꎬ 2021) ꎮ 调控ꎮ 然而ꎬ在紫色和白色花花冠对比中ꎬWRKY
苯丙氨酸是花青素合成的前体物质ꎬ经过一 和 ZIP 家族基因并未参与ꎬWRKY 与 ZIP 家族基因
系列酶及中间产物转化ꎬ最终在糖基转移酶的作 在紫色和白色花花冠对比中的缺失可能导致紫色
用下 生 成 有 色 花 青 素ꎮ 例 如ꎬ 在 牡 丹 ( Paeonia 表型的形成ꎮ 在矮牵牛( Petunia hybrida) 中ꎬPH3
suffruticosa)“二桥”粉白色和紫红色双色花的研究 是 AN11 ̄AN1 ̄PH4 复合体的目标基因ꎬ编码可与
中ꎬ两种花色差异基因主要富集于苯丙素和类黄 AN11 结 合 的 WRKY 蛋 白ꎬ 激 活 PH3 目 标 基 因
酮生物合成途径( Xu et al.ꎬ 2024)ꎮ 本研究中ꎬ紫 PH5ꎬ从而增加花青素含量( Verweij et al.ꎬ 2016)ꎻ
色和白色花花冠比较表明ꎬ苯丙素代谢途径的富 在火龙果(Hylocereus monacanthus)中ꎬHmoWRKY40
集可能在紫色花花冠的形成过程中可能发挥关键 可以结合并激活 HmoCYP76AD1 的启动子ꎬ升高甜
作用ꎮ 金属离子转运可能通过色素与金属离子缔 菜 色 素 含 量 ( Zhang et al.ꎬ 2021 )ꎻ 在 红 花
合形成色素-金属离子络合物ꎬ从而改变参与色素 (Carthamus tinctorius)中ꎬ属于 bZIP 家族的 PpHY5
生物合成、转化和破坏的酶的活性ꎬ进而调节花色 蛋白与 PpCHS1、PpCHS2 和 PpDFR 的 E ̄box 元件
(Li et al.ꎬ 2016)ꎮ 此外ꎬ颜色的变化可能不受单 结合ꎬ其表达的激活可导致花青素水平的增加( Li