
Page 93 - 《广西植物》2025年第1期
P. 93
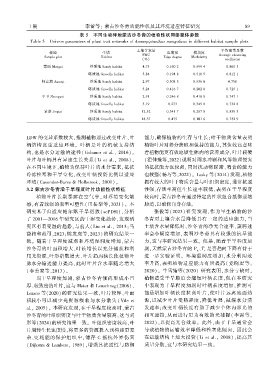
1 期 李蕾等: 蒙古沙冬青功能性状及其环境适应特征研究 8 9
表 5 不同生境样地蒙古沙冬青的植物性状网络整体参数
Table 5 Univers parameters of plant trait networks of Ammopiptanthus mongolicus in different habitat sample plots
土壤含水量 平均聚类系数
样地 生境 边密度 模块度
SWC Average clustering
Sample plot Habitat Edge degree Modularity
(%) coefficient
蒙西 Mengxi 沙质地 Sandy habitat 4.73 0.160 2 0.599 4 0.860 3
砾质地 Gravelly habitat 5.84 0.194 8 0.510 5 0.812 1
阿左旗 Azuoqi 沙质地 Sandy habitat 2.97 0.308 3 0.536 6 0.758
砾质地 Gravelly habitat 5.24 0.416 7 0.202 8 0.725 1
中卫 Zhongwei 沙质地 Sandy habitat 2.91 0.246 8 0.418 8 0.747 1
砾质地 Gravelly habitat 3.19 0.273 0.345 8 0.734 8
景泰 Jingtai 沙质地 Sandy habitat 12.52 0.341 7 0.287 8 0.859 3
砾质地 Gravelly habitat 18.37 0.475 0.101 6 0.754 9
LDW 的变异系数较大ꎬ推测植物通过改变叶片、叶 能力ꎬ确保植物的生存与生长ꎻ叶干物质含量表明
柄结构比重适应环境ꎮ 叶柄是叶的机械支持结 植物叶片对养分获取和保持的能力ꎬ其值较高意味
构ꎬ也是水分运输的途径( Gebauer et al.ꎬ 2016)ꎮ 着植物能更有效地锁住体内的营养成分ꎬ叶片较硬
叶片与叶柄具有异速生长关系( Li et al.ꎬ 2008)ꎬ (霍佳璇等ꎬ2022)说明对雨水冲刷和风等物理侵害
在不同生境下ꎬ植物为保持叶片的水分需求ꎬ抵抗 的抵抗能力也较强ꎬ同时抗动物踩踏、啃食的能力
冷冻栓塞和干旱空化ꎬ改变叶柄投资比例以适应 也较强(杨巧等ꎬ2022)ꎮ Lasky 等(2014)发现ꎬ植物
环境(Cavender ̄Bares & Holberookꎬ 2001)ꎮ 拥有较大的叶干物质含量与叶组织密度ꎬ通常抗逆
3.2 蒙古沙冬青沿干旱程度叶片功能性状特征 性强、存活率高但生长速率较低ꎬ表明在干旱程度
植物叶片长期暴露在空气中ꎬ对环境变化敏 较高时ꎬ蒙古沙冬青通过特定的性状组合抵御逆境
感ꎬ有着较强的表型可塑性( 田岳梨等ꎬ2021)ꎮ 本 胁迫ꎬ以确保自身存活ꎮ
研究基于自适应帕尔默干旱指数( scPDSI)ꎬ分析 张俊等(2023) 研究发现ꎬ作为旱生植物的沙
了 2001—2016 年研究区的干湿变化趋势ꎬ发现研 冬青对土壤含水量降低具有一定的适应能力ꎬ当
究区有着变湿的趋势ꎬ与前人(Liu et al.ꎬ 2013ꎻ马 土壤含水量降低时ꎬ沙冬青的净光合速率、蒸腾速
潇祎和范可ꎬ2023ꎻ陈发虎等ꎬ2023) 的研究结果一 率会小幅度增加ꎬ表明沙冬青具有较强的抗旱能
致ꎮ 随着干旱程度减弱和环境湿润度增加ꎬ蒙古 力ꎬ这与本研究结果一致ꎮ 但是ꎬ随着干旱程度加
沙冬青的叶面积增大、叶长增长以充分捕获和利 剧ꎬ天然蒙古沙冬青的 P 、T 是否趋向下降有待于
r
n
用光资源ꎬ叶形指数增大、叶片趋向狭长使植物叶 进一步实验证明ꎮ 环境湿润度增加ꎬ水分利用效
脉水分输送能力提高ꎬ此时叶片含水率随之增大 率升高ꎬ表明植物适应能力有所提高( 党晓宏等ꎬ
(李善家等ꎬ2013)ꎮ 2020)ꎮ 牛雪婧等(2020) 研究表明ꎬ水分亏缺时ꎬ
当干旱程度加剧ꎬ蒙古沙冬青倾向形成小且 植物遭受干旱胁迫会缩短叶柄长度ꎬ但在本研究
厚、较致密的叶片ꎬ这与 Meier 和 Leuschneg(2008)、 中表现为干旱程度加剧时叶柄长度增加ꎬ推测可
Lozano 等(2020) 的研究结果一致ꎬ叶片较厚、叶面 能是增加叶柄长度拉高叶片ꎬ使叶片远离地面热
积较小可以减少光资源吸收与水分散失 ( Vile et 源ꎬ以减少叶片受热程度ꎬ降低叶温ꎬ减缓水分蒸
al.ꎬ 2005)ꎮ 本研究发现ꎬ在干旱程度较高时ꎬ蒙古 发速率ꎻ改变叶柄长度有助于减少个体内部光的
沙冬青的叶组织密度与叶干物质含量较高ꎬ这与刘 相互遮挡ꎬ从而进行更为有效的光捕捉( 李露等ꎬ
彤等(2024)的研究结果一致ꎮ 叶组织密度较高ꎬ叶 2022)ꎬ以提高光合效率ꎮ 此外ꎬ由于干旱通常会
片周转生长速度慢ꎬ将更多的资源投入到构建更复 导致植物的运输效率降低和栓塞化倾向ꎬ 因此会
杂、更耗能的保护组织中ꎬ储存 C 抵抗外界伤害 在运输结构上加大投资( Li et al.ꎬ 2008)ꎬ提高其
(Dijkstra & Lambersꎬ 1989)ꎬ增强其抗逆性与防御 质量分数ꎬ这与本研究结果一致ꎮ