
Page 10 - 《广西植物》2025年第10期
P. 10
1 7 4 0 广 西 植 物 45 卷
体的质体小球数与总叶绿素含量呈显著负相关
(P = 0.037 4)ꎻ基粒数与总叶绿素含量呈显著正相
关(P = 0.016 7)ꎻ质体小球直径与叶绿素含量呈负
相关ꎬ但不显著ꎻ类胡萝卜素含量与质体小球数
量、直径及基粒数均无显著相关性(表 3)ꎮ
2.6 不同程度紫脉叶片中质体色素代谢及调控相
关基因的差异
对质体色素代谢相关基因( 类胡萝卜素合成
途径 关 键 酶 基 因) 及 色 素 代 谢 转 录 调 控 因 子
(AP2a、MYB12、PIF1a) 的 mRNA 水平变化进行了
检测(图 6)ꎮ 与无症组相比ꎬTSWV 侵染后不同紫
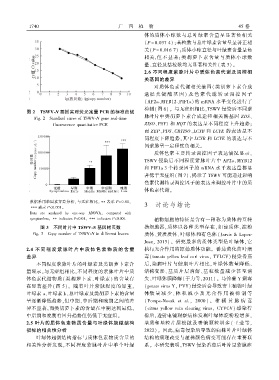
图 2 TSWV ̄N 基因实时荧光定量 PCR 的标准曲线
脉叶片中类胡萝卜素合成途径相关酶基因 ZDS、
Fig. 2 Standard curve of TSWV ̄N gene real ̄time
fluorescence quantitative PCR ZISO、PSY1 和 HQT 的表达呈不同程度上升趋势ꎻ
而 ZEP、PDS、CRTISO、LCYB 和 LCYE 的表达呈不
同程度下降趋势ꎬ其中 LCYB 和 LCYE 的表达与不
同紫脉呈一定程度负相关ꎮ
质体色素主要转录调控因子表达情况显示ꎬ
TSWV 侵染后不同程度紫脉叶片中 AP2a、MYB12
和 PIF1a 3 个转录因子的 mRNA 水平表达量都显
著低于无症组(图 7)ꎬ揭示了 TSWV 可能通过影响
色素代谢转录调控因子的表达来调控叶片中的质
体色素代谢ꎮ
数据采用单因素方差分析ꎬ与无症相比ꎬ∗∗表示 P<0.01ꎬ
3 讨论与结论
∗∗∗表示 P<0.001ꎮ
Data are analyzed by one ̄way ANOVAꎬ compared with
symptomlessꎬ ∗∗ indicates P<0.01ꎬ ∗∗∗ indicates P<0.001. 植物细胞的特征是含有一种称为质体的互转
图 3 不同叶片中 TSWV ̄N 基因拷贝数 换细胞器ꎬ质体以各种类型存在ꎬ如前质体、淀粉
Fig. 3 Copy number of TSWV ̄N in different leaves 质体、黄质质体、叶绿体和有色体( Jarvis & Lopez ̄
Juezꎬ 2013)ꎮ 研究最多的质体类型是叶绿体ꎬ它
2.4 不 同 程 度 紫 脉 叶 片 中 质 体 色 素 物 质 的 含 量 执行光合作用的原始质体功能ꎮ 番茄黄化曲叶病
差异 毒(tomato yellow leaf curl virusꎬ TYLCV) 侵染番茄
不同程度紫脉叶片的叶绿素及类胡萝卜素含 后ꎬ染病叶片与健康叶片相比ꎬ叶绿体数量降低、
量显示ꎬ与无症组相比ꎬ不同程度的紫脉叶片中质 结构变形、基质片层消解、基粒数量减少甚至消
体色素代谢物质( 类胡萝卜素、叶绿素) 的含量存 失、叶绿体膜降解(于力等ꎬ2011)ꎮ 马铃薯 Y 病毒
在显著差异( 图 5)ꎮ 随着叶片紫脉程度的加重ꎬ (potato virus Yꎬ PVY)侵染后会导致寄主植物叶绿
叶绿素 a、叶绿素 b、总叶绿素及类胡萝卜素的含量 体数 量 减 少、 体 积 减 小 及 光 合 作 用 被 抑 制 等
呈逐渐降低趋势ꎬ但中期、中后期和晚期之间的差 (Pompe ̄Novak et al.ꎬ 2000 )ꎮ 柑 橘 黄 脉 病 毒
异不显著ꎬ而类胡萝卜素的含量在中期达到最低ꎬ ( citrus yellow vein clearing virusꎬ CYVCV) 感染柠
中后期和晚期有回升趋势但仍低于无症组ꎮ 檬后ꎬ透射电镜观察病株发现叶绿体淀粉粒增多ꎬ
2.5 叶片的质体色素物质含量与叶绿体超微结构 基质和基粒片层松散及嗜锇颗粒增多( 王莹等ꎬ
指标的相关性分析 2022)ꎮ 因此ꎬ病毒侵染后导致的病株叶片叶绿体
叶绿体超微结构指标与质体色素物质含量的 结构的病理改变与植株颜色病变可能存在重要联
相关性分析发现ꎬ不同程度紫脉叶片中单个叶绿 系ꎮ 本研究表明ꎬTSWV 侵染番茄后叶片呈紫脉症