
Page 43 - 《广西植物》2025年第7期
P. 43
7 期 王莲哲等: 小麦 Tae ̄miR167 克隆及抗旱功能分析 1 2 3 5
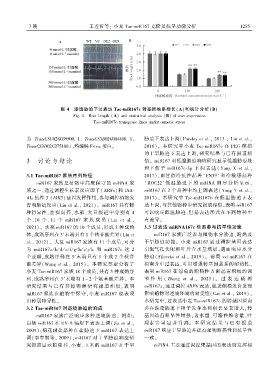
图 4 渗透胁迫下过表达 Tae ̄miR167c 转基因株系根长(A)和统计分析(B)
Fig. 4 Root length (A) and statistical analysis (B) of over ̄expression
Tae ̄miR167c transgenic lines under osmotic stress
为 TraesCSU02G039500. 1、 TraesCS3B02G004400. 1、 胁迫下表达上调( Pandey et al.ꎬ 2013 ꎻ Liu et al.ꎬ
TraesCS3D02G275100.1ꎬ均编码 F ̄box 蛋白ꎮ 2016)ꎮ 本研究中小麦 Tae ̄miR167c 在 PEG 模拟
的干旱胁迫下表达上调ꎬ研究结果与已有报道相
3 讨论与结论 似ꎮ miR167 对低温胁迫响应研究显示低温胁迫处
理下茄 子 miR167c ̄3p 下 调 表 达 ( Yang X et al.ꎬ
3.1 Tae ̄miR167 家族序列特征 2017)ꎬ而甘蔗冷抗性品种‘ FN39’ 和冷敏感品种
miR167 家族是植物中高度保守的 miRNA 家 ‘ROC22’ 低 温 胁 迫 下 的 miRNA 测 序 分 析 显 示ꎬ
族之一ꎬ通过调控生长素反应因子( ARFs) 和 IAA ̄ miR167 在 2 个品种中均上调表达( Yang Y et al.ꎬ
Ala 抗性 3 (IAR3)基因发挥作用ꎬ参与调控植物发 2017)ꎮ 本研究中 Tae ̄miR167b 在低温胁迫下表
育和胁迫反应( Liu et al.ꎬ 2021)ꎮ miR167 具有物 达上调ꎬ与其他物种中研究报道相似ꎬ表明 miR167
种特异性ꎬ在拟南芥、水稻、大豆报道中分别有 4 可以响应低温胁迫ꎬ但是表达形式在不同物种中
个、10 个、 11 个 miR167 家 族 成 员 ( Liu et al.ꎬ 有差异ꎮ
2021)ꎮ 水稻 miR167 的 10 个成员ꎬ形成 2 种成熟 3.3 过表达 miRNA167c 株系参与抗旱性功能
体ꎬ成熟序列在 3′末端只有 1 个核苷酸差异(Liu et miR167 家族广泛参与植物水分胁迫、耐热及
al.ꎬ 2012)ꎮ 大豆 miR167 家族有 11 个成员ꎬ可分 干旱胁迫功能ꎮ 小麦 miR167 通过调控基因表达
为 miR167a / b / d / e / f/ g / h / i/ j/ k 和 miR167c 这 2 引起气孔关闭和叶片含水量增加ꎬ进而响应水分
个亚群ꎬ成熟序列在 3′末端只有 1 个或 2 个核苷 胁迫( Fileccia et al.ꎬ 2019)ꎮ 葡萄 vvi ̄miR167 在
酸差异(Wang et al.ꎬ 2015)ꎮ 本研究鉴定分析了 拟南芥中过表达ꎬ可以增强转基因品系的耐热性ꎬ
小麦 Tae ̄miR167 家族 18 个成员ꎬ共有 3 种成熟序 表明 miR167 在葡萄的耐热性方面起着积极的调
列ꎬ成熟序列在 3′末端有 1 ~ 2 个氨基酸差异ꎮ 本 节 作 用 ( Zhang et al.ꎬ 2023 )ꎮ 过 表 达 杨 树
研究结 果 与 已 有 其 他 物 种 研 究 报 道 相 似ꎬ 表 明 miR167aꎬ通过调控 ARF8 表达ꎬ促进侧根发育进而
miR167 家族在植物中保守ꎬ小麦 miR167 也表现 影响植物对逆境环境的耐受性( Cai et al.ꎬ 2019)ꎮ
出种属特异性ꎮ 本研究中ꎬ过表达小麦 Tae ̄miR167c 的转基因拟南
3.2 Tae ̄miR167 对逆境胁迫的响应 芥在渗透胁迫下种子发芽率和根长显著增大ꎬ转
miR167 家族广泛响应多种逆境胁迫ꎮ 例如: 基因幼苗耐旱性增强ꎬ含水量、可溶性糖含量、叶
白杨 miR167 在 UV ̄B 辐射下表达上调( Jia et al.ꎬ 绿素含 量 显 著 升 高ꎮ 本 研 究 结 果 与 已 有 报 道
2009)ꎻ棉花耐盐品种在盐胁迫下 miR167 表达上 miR167 响应干旱胁迫并提高植物耐热性和抗旱性
调(李春贺等ꎬ 2009)ꎻmiR167 对干旱胁迫响应研 一致ꎮ
究报道显示拟南芥、小麦、玉米的 miR167 在干旱 miRNA 主要通过调控靶基因的表达而发挥相