
Page 123 - 《广西植物》2026年第2期
P. 123
2 期 康瑶瑶等: 滇西北水蓼叶功能性状的关联性及其对气候因子的响应 3 1 7
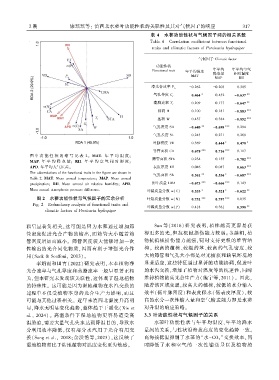
表 4 水蓼功能性状与气候因子间的相关系数
Table 4 Correlation coefficient between functional
traits and climatic factors of Persicaria hydropiper
气候因子 Climatic factor
功能性状
年平均 年平均空气
Functional trait 年平均温度
降水量 相对湿度
MAT
MAP RH
净光合速率 P n -0.262 -0.203 0.305
0.464 ∗ 0.432 -0.637 ∗∗
气孔导度 G s
蒸腾速率 T r 0.209 0.177 -0.647 ∗∗
株高 H -0.100 -0.181 -0.883 ∗∗∗
茎粗 W 0.437 0.384 -0.852 ∗∗∗
气孔密度 SD -0.640 ∗∗ -0.698 ∗∗∗ -0.294
气孔长度 SL 0.243 0.271 0.268
叶脉密度 VD 0.369 0.444 ∗ 0.478 ∗
导管面积 CA 0.678 ∗∗∗ 0.716 ∗∗∗ -0.117
图 中 功 能 性 状 的 缩 写 见 表 2ꎮ MAT. 年 平 均 温 度ꎻ
筛管面积 STA 0.234 0.155 -0.782 ∗∗∗
MAP. 年 平 均 降 水 量ꎻ RH. 年 平 均 空 气 相 对 湿 度ꎻ
APD. 年平均大气压差ꎮ 表皮厚度 ET 0.046 0.087 0.663 ∗∗∗
The abbreviations of the functional traits in the figure are shown in
气孔面积 SA 0.561 ∗∗ 0.536 ∗ -0.687 ∗∗∗
Table 2. MAT. Mean annual temperatureꎻ MAP. Mean annual
precipitationꎻ RH. Mean annual air relative humidityꎻ APD. 比叶质量 LMA -0.673 ∗∗∗ -0.666 ∗∗∗ -0.143
Mean annual atmospheric pressure difference.
叶碳质量分数 ω(C) 0.533 ∗ 0.521 ∗ -0.622 ∗∗
图 2 水蓼功能性状与气候因子的冗余分析 叶氮质量分数 ω(N) 0.751 ∗∗∗ 0.787 ∗∗∗ -0.035
Fig. 2 Redundancy analysis of functional traits and
叶磷质量分数 ω(P) -0.418 -0.361 0.598 ∗∗
climatic factors of Persicaria hydropiper
积呈显著负相关ꎬ这可能是因为水蓼通过增加筛 Sun 等(2016) 研究表明ꎬ植株越高更容易获
管密度促进光合产物的输出ꎬ而筛管大小随着筛 得更多的光ꎬ但抗机械损伤能力较弱ꎻ茎越粗ꎬ植
管密度增加而减小ꎮ 筛管密度较大能够增加一次 物抗机械损伤能力越强ꎬ同时支持更粗的维管结
性输出的光合同化物质ꎬ因而有利于增强光合作 构ꎮ 较高的植株、较粗的茎、较高的气孔密度、较
大的筛管和气孔大小都是对光捕获和缺氧环境的
用(Sack & Scoffoniꎬ 2013)ꎮ
李娟霞和田青(2022) 研究表明ꎬ木本植物净 显著适应ꎬ这些特征通过显著的功能协调ꎬ促进叶
光合速率与气孔导度和蒸腾速率一般呈显著正相 的水汽交换ꎬ增加了植物对温度等的抗逆性ꎬ同时
关ꎬ但本研究未发现该关联性ꎬ这体现了湿地植物 维持植物的高光合生产力( 施宇等ꎬ2011)ꎮ 因此ꎬ
的特殊性ꎮ 这可能是因为湿地植物在水汽交换的 随着该区域变暖ꎬ较高大的植株ꎬ较低的水分输入
过程中不仅受植物本身的光合生产力影响ꎬ而且 效率(低叶脉密度) 和表皮保水( 低表皮厚度)ꎬ较
可能与其他过程相关ꎮ 近年来滇西北温度升高明 高的水分一次性输入量和空气输送能力即是水蓼
显ꎬ降水无明显变化趋势ꎬ整体趋于干暖化(Xie et 对升温的响应策略ꎮ
al.ꎬ 2024)ꎮ 高温条件下湿地植物更容易遭受高 3.3 叶功能性状与气候因子的关系
温胁迫ꎬ需要大量气孔失水达到降温目的ꎬ导致水 水蓼叶功能性状与年平均温度、年平均降水
分利用效率降低ꎬ仅有部分水汽用于光合有用交 量间的关系ꎬ与性状沿海拔高度的变化趋势一致ꎮ
换(Song et al.ꎬ 2018ꎻ余洪艳等ꎬ2023)ꎬ这反映了 高海拔低温抑制了水蓼的“ 水-CO ” 交换效率ꎬ同
2
湿地植物相比于陆地植物对温度变化更为敏感ꎮ 时降低了水和空气的一次性输送量以及植物的