
Page 26 - 《广西植物》2021年第12期
P. 26
ϟ ँ Ꭾ ԡ 广জ 西জ 植জ 物 ࿗ϟ 卷
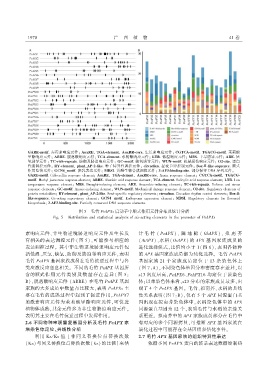
ᤩዛቴጛቂᅳᡙᔠஏ 赤霉素响应元件ଫ ዛࣩଝቴጛᖔ ௴ᤩዛᓣᤀᓣቂᓣႥᡙᖔ ዛࣩଝቴቴዞᅳʗᓣ 生长素响应元件ଫ ࣷᤩ௴ࣷዛቂᅳᡙᔠஏᖔ ௴ᤩዛࣷᤩቂᅳᡙᔠஏ 茉莉酸
甲酯响应元件ଫ ዛଧቴጛ 脱落酸响应元件ଫ ௴ࣷዛᓣᤀᓣቂᓣႥᡙ 水杨酸响应元件ଫ ᒦ௴ቴ 低温响应元件ଫ ᤹ଧ 干旱诱导元件ଫ ዛቴጛ 厌
氧诱导元件ଫ ௴ࣷʗᔠዞ၊ʗᓣᡓᓣᐹᡙ 防御及胁迫响应元件ଫ ᤩࣷቂᅳᡙᔠஏ 缺氧诱导元件ଫ ۩Іટቂᅳᡙᔠஏ 机械损伤响应元件ଫ ᧣ԣᔠᡙᓣ 蛋白
代谢调控元件ଫ ቴɔᓣᤀᓣቂᓣႥᡙᖔ ᡓᤀᐹႥᡙᡚዛۗԣᤀᔠҧᓣ 种子特异性调控元件ଫ ዞᔠʗዞᐹʐᔠᐹႥ 昼夜节律控制元件ଫ ଧᅳଝտᤀᔠҧᓣᓣफࣩᓣႥዞᓣ 顺式
作用监管元件ଫ ᤩࣷટᡚቂᅳᡙᔠஏ 胚乳表达元件ଫ ᤹ଧᧇ 黄酮生物合成调控元件ଫ ᇺዛÞᇺᤇᔠႥʐᔠႥᔠᡙᓣ 部分保守 ᧕શዶ 序列元件ᤥ
ᤩዛቴጛቂᅳᡙᔠஏ ᥈ᔽᤦᤦᔀʢᔀᔽႿ ʢᔀ༁ᡱᆍႿ༁ᔀ ᔀᔀቝᔀႿᡷଫ ዛࣩଝቴጛᖔ ௴ᤩዛᓣᤀᓣቂᓣႥᡙᖔ ዛࣩଝቴቴዞᅳʗᓣ ዶࣼᔽႿ ʢᔀ༁ᡱᆍႿ༁ᔀ ᔀᔀቝᔀႿᡷଫ ࣷᤩ௴ࣷዛቂᅳᡙᔠஏᖔ ௴ᤩዛࣷᤩ
ቂᅳᡙᔠஏ ᥘᔀᡷၤᢃ ᢼᑕ༁ቝᆍႿᑕᡷᔀ ʢᔀ༁ᡱᆍႿ༁ᔀ ᔀᔀቝᔀႿᡷଫ ዛଧቴጛ ዶᤦ༁ዹᔽ༁ᔽዹ ᑕዹᔽʛ ʢᔀ༁ᡱᆍႿ༁ᔀ ᔀᔀቝᔀႿᡷଫ ௴ࣷዛᓣᤀᓣቂᓣႥᡙ ୩ᑕᔽዹᢃᔽዹ ᑕዹᔽʛ ʢᔀ༁ᡱᆍႿ༁ᔀ ᔀᔀቝᔀႿᡷଫ ᒦ௴ቴᓂᆍऔ
ᡷᔀቝᡱᔀʢᑕᡷࣼʢᔀ ʢᔀ༁ᡱᆍႿ༁ᔀ ᔀᔀቝᔀႿᡷଫ ᤹ଧ ᧕ʢᆍࣼ੫ၤᡷ᥋ᔽႿʛࣼዹᔽႿ੫ ᔀᔀቝᔀႿᡷଫ ዛቴጛ ዶႿᑕᔀʢᆍᤦᔽዹ᥋ᔽႿʛࣼዹᔽႿ੫ ᔀᔀቝᔀႿᡷଫ ௴ࣷʗᔠዞ၊ʗᓣᡓᓣᐹᡙ ᧕ᔀᔀႿ༁ᔀ ᑕႿʛ ༁ᡷʢᔀ༁༁
ʢᔀ༁ᡱᆍႿ༁ᔀ ᔀᔀቝᔀႿᡷ༁ଫ ᤩࣷቂᅳᡙᔠஏ ዶႿᆍᔽዹ᥋ᔽႿʛࣼዹᔽႿ੫ ᔀᔀቝᔀႿᡷଫ ۩Іટቂᅳᡙᔠஏ ᥘᔀዹၤᑕႿᔽዹᑕ ʛᑕቝᑕ੫ᔀ ʢᔀ༁ᡱᆍႿ༁ᔀ ᔀᔀቝᔀႿᡷଫ ᧣ԣᔠᡙᓣ ᔀ੫ࣼᑕᡷᆍʢᢃ ᔀᔀቝᔀႿᡷ༁ ᆍ
ᡱʢᆍᡷᔀᔽႿ ቝᔀᡷᑕᤦᆍᔽ༁ቝଫ ቴɔᓣᤀᓣቂᓣႥᡙᖔ ᡓᤀᐹႥᡙᡚዛۗԣᤀᔠҧᓣ ୩ᔀᔀʛ᥋༁ᡱᔀዹᔽᔽዹ ʢᔀ੫ࣼᑕᡷᆍʢᢃ ᔀᔀቝᔀႿᡷ༁ଫ ዞᔠʗዞᐹʐᔠᐹႥ ऊᔽʢዹᑕʛᔽᑕႿ ʢၤᢃᡷၤቝ ዹᆍႿᡷʢᆍ ᔀᔀቝᔀႿᡷ༁ଫ ଧᅳଝտ
ᤀᔠҧᓣᓣफࣩᓣႥዞᓣ ࣷᔠ᥋ᑕዹᡷᔽႿ੫ ༁ࣼᡱᔀʢᔽ༁ᆍʢᢃ ᔀᔀቝᔀႿᡷଫ ᤩࣷટ ᡚ ቂᅳᡙᔠஏ ጶႿʛᆍ༁ᡱᔀʢቝ ᔀᡱʢᔀ༁༁ᔽᆍႿ ᔀᔀቝᔀႿᡷଫ ᤹ଧᧇ ᔀ੫ࣼᑕᡷᆍʢᢃ ᔀᔀቝᔀႿᡷ༁ ᆍʢ ᑕᆍႿᆍᔽʛ
ᤦᔽᆍ༁ᢃႿᡷၤᔀ༁ᔽ༁ଫ ᇺዛÞᇺᤇᔠႥʐᔠႥᔠᡙᓣ ۪ᑕʢᡷᔽᑕᢃ ዹᆍႿ༁ᔀʢᔀʛ ᧕શዶ ༁ᔀीࣼᔀႿዹᔀ ᔀᔀቝᔀႿᡷ༁
图 ጢজ 毛竹 ۗᓣዛۗᣏ 启动子中顺式作用元件分布及统计分析
èᔽ੫ ጢজ ᧕ᔽ༁ᡷʢᔽᤦࣼᡷᔽᆍႿ ᑕႿʛ ༁ᡷᑕᡷᔽ༁ᡷᔽዹᑕ ᑕႿᑕᢃ༁ᔽ༁ ᆍ ዞᔠ᥋ᑕዹᡷᔽႿ੫ ᔀᔀቝᔀႿᡷ༁ ᔽႿ ᡷၤᔀ ᡱʢᆍቝᆍᡷᔀʢ ᆍ ۗᓣዛۗᣏ
素响应元件ܦ非生物逆境胁迫响应元件及生长发 计毛 竹 ྉ ۗᓣዛۗᣏ ɯܦ 陆地棉 ྉ ᤩ၊ዛۗᣏ ɯܦ 拟南芥
育相关的表达调控元件ྉ 图 ጢɯᖔ可能参与相应的 ྉዛᡙዛۗᣏɯܦ水稻ྉ ᧣ዛۗᣏɯ 的 ዛۗᣏ 基因家族成员的
表达调控过程ᤥ 其中非生物逆境胁迫响应元件包 进化选择模式ᖔ比值均小于 ϟྉ 图 ᤃɯᖔ表明各物种
括低温ܦ厌氧ܦ缺氧ܦ防御及胁迫等响应元件ᖔ表明 的 ዛۗᣏ 基因家族成员都为纯化选择ᤥ 毛竹 ۗᓣዛۗᣏ
毛竹 ۗᓣዛۗᣏ 基因家族成员在毛竹抗逆过程中与各 基因家族 ϟ 个家族成员定位于 ϟሕ 条染色体上
类应激反应息息相关ᤥ 不同的毛竹 ۗᓣዛۗᣏ 基因所 ྉ图 Ꭾ᧥ዶɯᖔ不同染色体基因分布密度存在差异ᖔ以
含的顺式作用元件类别及数量存在差异 ྉ 图 ጢ᧥ ༁ϟሕ 密度居高ᖔۗᓣዛۗᣏᤃܦۗᓣዛۗᣏϟᤃ 均定位于该染色
ାɯᖔ脱落酸响应元件 ྉዶାጶɯ 在毛竹 ۗᓣዛۗᣏ 基因 体ଫ以单染色体来看ᖔ༁ሕ 分布的家族成员最多ᖔ出
家族的大多成员中数量占比较大ᖔ表明 ۗᓣዛۗᣏ 主 现了 ࿗ 个 ۗᓣዛۗᣏ 基因ᤥ 毛竹ܦ拟南芥ܦ水稻的共线
要在毛竹的成熟过程中起到了促进作用ᖔۗᓣዛۗᣏᎮ 性关系表明ྉ 图 Ꭾ᧥ାɯᖔ仅有 ጢ 个 ዛۗᣏ 同源蛋白基
的激素响应元件为茉莉酸甲酯响应元件ᖔ可促进 因出现在拟南芥染色体中ᖔ水稻染色体中的 ዛۗᣏ
植物体成熟ᖔ其余元件多为非生物胁迫响应元件ᖔ 同源蛋白基因为 ϟ 个ᖔ表明毛竹与水稻的亲缘关
表明其主要在毛竹抗逆过程中发挥作用ᤥ 系更近ᤥ 拟南芥中的 ዛۗᣏ 家族成员部分在毛竹中
ԣॹᣤ 不同物种串联重复基因分析及毛竹 ۗᓣዛۗᣏ 家 有对应的多个同源拷贝ᖔ可推断 ዛۗᣏ 基因家族在
族染色体定位ܓ共线性分析 演化过程中可能存在全基因组多倍化事件ᤥ
利用 ᑕ ᣰ ༁ 值 Џ 非 同 义 替 换 位点替换次数 ԣॹ᎒ 毛竹 ዛۗᣏ 基因家族的组织特异性表达
ྉᑕɯ与同义替换位点替换次数ྉ༁ɯ 的比值 来估 依据不同 ۪ᔀዶ۪ᣮ 蛋白的转录表达数据绘制热