
Page 29 - 《广西植物》2020年第4期
P. 29
4 期 刘成等: 基于 SRAP 标记的紫溪山华山松种子园无性系遗传多样性分析 4 6 7
种源单独聚为一类ꎮ
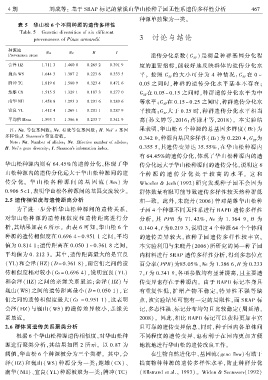
表 5 华山松 6 个不同种源的遗传多样性
Table 5 Genetic diversities of six different
provenances of Pinus armandii 3 讨论与结论
种源地
Na Ne H I
Provenance areas 遗传分化系数( G ) 是衡量种群基因分化程
ST
会泽 HZ 1.711 3 1.460 0 0.265 2 0.391 9 度的重要指标ꎬ能较好地反映群体的遗传分化水
巍山 WS 1.644 3 1.387 2 0.223 6 0.333 5 平ꎮ 按照 G 的大小可分为 4 种 情 况:G 在 0 ~
ST
ST
腾冲 TC 1.819 6 1.580 9 0.323 4 0.471 6 0.05 之间时ꎬ种群的遗传分化水平基本不存在ꎻ
楚雄 CX 1.515 5 1.329 1 0.187 3 0.277 0 G 在 0.05 ~ 0.15 之间时ꎬ种群遗传分化水平为中
ST
南华 NH 1.458 8 1.293 3 0.201 6 0.289 6 等水平ꎻG 在 0.15 ~ 0.25 之间时ꎬ种群遗传分化水
ST
宜良 YL 1.412 4 1.269 1 0.201 1 0.287 9 平较高ꎻG 大于 0.25 时ꎬ种群遗传分化水平相当
ST
平均值 Mean 1.593 7 1.386 6 0.233 7 0.341 9 高(孙文婷等ꎬ2016ꎻ蒋谦才等ꎬ2018)ꎮ 本实验结
注: Na. 等位基因数ꎻ Ne. 有效等位基因数ꎻ H. Nei′ s 基因 果表明ꎬ华山松 6 个种源的总基因多样度( Ht) 为
多样性ꎻI. Shannons 信息指数ꎮ 0.342 0ꎬ种源内基因多样性( Hs) 为 0.220 4ꎬG 为
Note: Na. Number of allelesꎻ Ne. Effective number of allelesꎻ ST
0.355 5ꎬ其遗传变异达 35.55%ꎬ在华山松种源内
H. Nei′s gene diversityꎻ I. Shannons information index.
有 64.45%的遗传分化ꎬ体现了华山松种源内的遗
华山松种源内则有 64.45%的遗传分化ꎬ体现了华 传分化远大于华山松种源间的遗传分化ꎬ说明这 6
山松种源内的遗传分化远大于华山松种源间的遗 个种 源 的 遗 传 分 化 处 于 较 高 的 水 平ꎮ 这 和
传分 化ꎮ 华 山 松 各 种 源 间 的 基 因 流 ( Nm) 为 Wheeler & Jech(1992) 研究发现种子园不会因为
0.906 5<1ꎬ表明华山松各种源间的基因交流较少ꎮ 群体数量有限可能导致遗传多样性较天然种群低
2.5 遗传相似度与遗传距离分析 相一致ꎮ 此外ꎬ朱晓丹(2006) 曾对楚雄华山松种
为了进一步分析华山松种源间的遗传关系ꎬ 子园 4 个种源不同无性系进行 RAPD 遗传多样性
对华山松种源的遗传相似度和遗传距离进行分 分析ꎬ 其 PPB 为 71. 42%ꎬ Ne 为 1. 364 9ꎬ H 为
析ꎬ其结果如表 6 所示ꎮ 由表 6 可知ꎬ华山松 6 个 0.140 4ꎬI 为0.219 3ꎬ说明这 4 个种源 64 个个体间
种源的遗传相似度在 0.696 4 ~ 0.951 1 之间ꎬ平均 的遗传差异较大ꎬ该种子园遗传多样性较丰富ꎮ
值为 0.814 1ꎻ遗传距离在 0.050 1 ~ 0.361 8 之间ꎬ 本实验利用与朱晓丹(2006)所研究的同一种子园
平均值为 0. 212 3ꎮ 其中ꎬ遗传距离最大的是宜良 内材料进行 SRAP 遗传多样性分析ꎬ得到多态位点
(YL)和会泽(HZ)(D = 0.361 8)ꎬ而它们之间的遗 百分率(PPB)为85.05%ꎬNe 为 1.386 6ꎬH 为 0.233
传相似度相对较小(Gs = 0.696 4)ꎬ说明宜良( YL) 7ꎬI 为 0.341 9ꎬ各项参数均有显著提高ꎬ且主要遗
和会泽( HZ) 之间的亲缘关系最远ꎻ会泽( HZ) 与 传变异也存在于种源内ꎮ 由于 RAPD 标记本身具
巍山(WS)之间的遗传距离最小( D = 0.050 1)ꎬ它 有重复性低ꎬ扩增产物不稳定ꎬ特异性不强等缺
们之间的遗传相似度最大(Gs = 0.951 1)ꎬ这表明 点ꎬ该实验结果可能有一定的局限性ꎬ而 SRAP 标
会泽(HZ)与巍山( WS) 的遗传差异较小ꎬ亲缘关 记ꎬ多态性强、标记分布均匀且比较稳定( 周延清ꎬ
系最近ꎮ 2008)ꎮ 因此ꎬ相比 RAPD 标记可以获得更加丰富
2.6 群体间遗传关系聚类分析 且可靠的遗传变异信息ꎬ同时ꎬ种子园内各单株间
根据 6 个华山松种源遗传相似度ꎬ对华山松种 不同程度的遗传变异ꎬ也有利于在园内更加方便
源进行聚类分析ꎬ其结果如图 2 所示ꎮ 以 0.87 为 地就地进行华山松的遗传改良工作ꎮ
阈值ꎬ华山松 6 个种源被分为三个类群ꎮ 其中ꎬ会 在生物自然进化中ꎬ基因流(gene flow)有助于
泽(HZ)和巍山( WS) 种源分为一类ꎻ楚雄( CX)、 提高物种种源的遗传多样性水平ꎬ防止种群分化
南华(NH)、宜良(YL)种源被聚为一类ꎻ腾冲(TC) (Ellstrand et al.ꎬ 1993)ꎮ Widen & Svensson(1992)