
Page 184 - 《广西植物》2022年第12期
P. 184
2 1 7 4 广 西 植 物 42 卷
究在红蓝光质基础上添加少量绿光与紫光后ꎬ显
著提高了 12 × 8R1B1P1G 和 16 × 6R1B1P1G 处理
的 叶 绿 素 和 类 胡 萝 卜 素 含 量ꎬ 降 低 了 12 ×
6R1B1P1G 处理的叶绿素和类胡萝卜素含量ꎬ这
可能是红蓝组合光基础上添加其他光质导致了互
补效应的产生ꎬ进而导致植物光合色素的合成与
积累的不同ꎮ 较 12 hd 光周期ꎬ16 hd 光周
 ̄1
 ̄1
期处理促进香子含笑叶片叶绿素和类胡萝卜素含
量和叶绿素 a / b 比值增加ꎬ能更有效地促进叶片
对光能的转换、捕获和传递能力ꎬXu 等(2020) 在
杉木组培苗中也得出相似的研究结果ꎮ
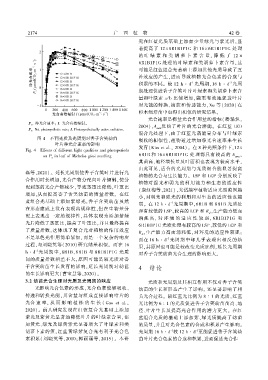
光合速率是衡量光合作用量的指标(潘瑞炽ꎬ
P n . 净光合速率ꎻ I. 光合有效辐射ꎮ
2004)ꎬA 反映了叶片的光合潜能ꎮ 在红蓝 LED
P n . Net photosynthetic rateꎻ I. Photosynthetically active radiation. max
混合光处理下ꎬ由于红蓝光谱能量分布与叶绿素
图 4 不同光质及光周期对香子含笑幼苗 吸收的相似性ꎬ植物通过增加净光合速率来生长
叶片净光合速率的影响
发育(Kim et al.ꎬ 2004)ꎮ 在 8 种光照条件下ꎬ12×
Fig. 4 Effects of different light qualities and photoperiods
6R1B 和 16 × 6R1B1P1G 处理都具有较高的 A ꎬ
on P in leaf of Michelia gioii seedling
n max
其苗高、地径增长量及叶面积也表现为较高水平ꎬ
由此可见ꎬ适合的光周期与光质组合能显著提高
磊等ꎬ2021)ꎮ 延长光周期使香子含笑叶片进行光
植物的光合与生长能力ꎮ LSP 和 LCP 分别反映了
合作用时长增加ꎬ光合产物分配向叶片倾斜ꎬ使分
植物对强光和弱光的利用能力和生态的适应性
配到茎的光合产物减少ꎬ导致茎重比降低ꎬ叶重比
(康红梅等ꎬ2021)ꎬ光谱越窄植物适应光照范围越
增加ꎬ从而提高香子含笑幼苗的质量指数ꎮ 在红
小ꎬ对弱光和强光的利用和对生态的适应性也越
蓝组合光基础上添加紫绿光ꎬ香子含笑幼苗虽然
弱ꎮ 在 12 hd 光周期中ꎬ6R1B 和 8R1B 光质处
 ̄1
在形态建成上没有表现出规律性ꎬ但在生物量分
理有较低的 LSPꎬ较高的 LCP 和 R ꎬ生产能力低而
d
配上表现出一定的规律性ꎬ具体表现为添加紫绿
消耗 高ꎬ 对 环 境 的 适 应 性 较 弱ꎬ 6R1B1P1G 和
光后降低了茎重比ꎬ提高了叶重比ꎬ并且最终提高
6R1B1P1G 光质处理有较高的 LSPꎬ较低的 LCP 和
了质量指数ꎬ这体现了复合光对植物的作用效应
R ꎬ生产能力高而消耗低ꎬ对环境的适应性较强ꎻ
d
不是单色光作用简单累加ꎬ而是一个复杂的响应
而在 16 hd 光周期中却几乎表现出相反的结
 ̄1
过程ꎬ与刘晓英等(2010) 研究结果相似ꎮ 而在 16
果ꎬ其原因也可能是较改变光质比例ꎬ延长光周期
 ̄1
hd 光周期中ꎬ8R1B、6R1B 与 8R1B1P1G 光质
对香子含笑幼苗光合生理的影响更大ꎮ
间的质量指数相差不大ꎬ原因可能是较光质对香
子含笑幼苗生长发育的影响ꎬ延长光周期对幼苗 4 结论
的生长影响更大(曹宝慧等ꎬ2020)ꎮ
3.2 幼苗光合生理对光质及光周期的响应 光质和光周期及其相互作用不仅对香子含笑
光影响光合色素的形成ꎬ光合色素能够吸收、 幼苗的生长和形态产生了影响ꎬ 还显著影响了相
传递和转换光能ꎬ其含量与组成直接影响叶片的 关光合过程ꎮ 较红蓝光比例为 8 ∶ 1 的光质ꎬ红蓝
光合 速 率ꎬ 从 而 影 响 植 株 的 生 长 ( Gao et al.ꎬ 光比例为 6 ∶ 1 的光质促进香子含笑幼苗苗高、地
2020)ꎮ 前人研究发现在红蓝复合光基础上添加 径、叶片生长及提高光合作用的潜力更大ꎮ 在红
紫光及紫黄光显著地降低叶片的叶绿素含量ꎬ添 蓝组合光质的基础上添加紫、绿光质提高了幼苗
加黄光、绿光及绿黄紫光显著增大了叶绿素和类 的质量ꎬ并且对光合色素的合成和积累产生影响ꎮ
胡萝卜素的值ꎬ红蓝黄绿紫复合光有利于光合色 光周期 16 hd 较 12 hd 更能促进香子含笑幼
 ̄1
 ̄1
素积累( 刘晓英等ꎬ2010ꎻ郭丽丽等ꎬ2015)ꎮ 本研 苗叶片光合色素的合成和积累ꎬ进而促进光合作