
Page 70 - 《广西植物》2023年第9期
P. 70
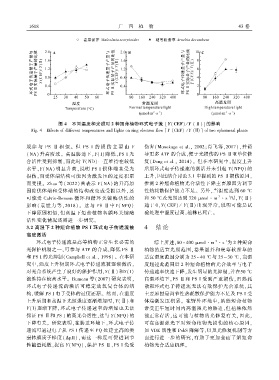
1 6 1 8 广 西 植 物 43 卷
图 4 不同温度和光照对 2 种短命植物环式电子流 [Y(CEF) / Y (Ⅱ)]的影响
Fig. 4 Effects of different temperatures and lights on ring electron flow [Y (CEF) / Y (Ⅱ)] of two ephemeral plants
规 律 与 PS Ⅱ 相 似ꎮ 但 PS I 的 损 伤 主 要 由 Y 伤害(Munekage et al.ꎬ 2002ꎻ肖飞等ꎬ2017)ꎬ并诱
( NA) 升高所致ꎮ 高温胁迫下ꎬY( I) 降低ꎬPS I 光 导更多 ATP 的合成ꎬ便于光损伤的 PS Ⅱ亚单位修
合活性受到抑制ꎬ而此时 Y( ND) 一直维持在较低 复(Deng et al.ꎬ 2014)ꎮ 但在本研究中ꎬ温度上升
水平ꎬY( NA) 明显升高ꎬ说明 PS I 供体端未受光 后期环式电子传递流的激活并未引起 Y( NPQ) 的
损伤ꎬ而受体端结构可能因为激发压的过度积累 上升ꎬ因此结合讨论 3.1 中提到的 PS Ⅱ损伤原因ꎬ
而受损ꎮ Zhao 等(2022) 曾表示 Y( NA) 的升高原 推测 2 种短命植物光合活性下降主要原因为调节
因除供体端和受体端的结构改变或受损以外ꎬ还 性热耗散保护能力不足ꎮ 另外ꎬ当温度达到 60 ℃
可能受 Calvin ̄Benson 循环和循环关键酶活性的 和 50 ℃ 及光照达到 320 μmolm s 时ꎬY( Ⅱ)
 ̄2
 ̄1
影响( 袁世力等ꎬ2018) ꎮ 这与 PS Ⅱ中Y( NPQ) 趋于 0ꎬY( CEF) / Y(Ⅱ) 出现异常ꎬ说明可能是试
下降原因相似ꎬ但高温下短命植物各循环关键酶 验处理中温度过高ꎬ植株已死亡ꎮ
活性变化情况还需进一步研究ꎮ
3.2 高温下 2 种短命植物 PS I 环式电子传递流被 4 结论
高度激活
环式电子传递流是高等植物正常生长必需的 综上所述ꎬ80 ~ 400 μmolm s 为 2 种短命
 ̄2
 ̄1
光保护机制之一ꎬ可参与 ATP 的合成ꎬ降低 PS Ⅱ 植物的适宜光照范围ꎬ卷果涩芥和硬萼软紫草的
和 PS I 的光抑制(Campbell et al.ꎬ 1998)ꎮ 在本研 适宜温度范围分别为 25 ~ 40 ℃ 与 25 ~ 30 ℃ ꎬ当温
究中ꎬ温度上升初期环式电子传递流被逐渐激活ꎬ 度超过此范围后 2 种短命植物的光合效率与电子
对光合系统产生了良好的保护作用ꎬY(Ⅱ)和Y(I) 传递速率快速下降ꎬ发生明显的光抑制ꎬ并在50 ℃
被维持在较高水平ꎮ Rumeau 等(2007) 研究表明ꎬ 高温环境下ꎬPS Ⅱ和 PS I 受到严重损伤ꎬ且热耗
环式电子传递流的激活可稳定放氧复合体的结 散和环式电子传递流无法有效保护光合系统ꎬ其
构ꎬ缓解 PS I 电子受体的过度还原ꎮ 然而ꎬ在温度 主要原因是调节性热耗散保护能力不足及 PS I 受
上升后期和高温下光照强度逐渐增加时ꎬY( Ⅱ) 和 体端激发压积累ꎮ 在野外环境中ꎬ虽然短命植物
Y(I)逐渐下降ꎬ环式电子传递速率的增加也无法 常受正午短时间内高温强光的胁迫ꎬ但植株依然
保证 PS Ⅱ和 PS I 的高光合活性ꎬ这与 Y(NPQ) 的 能正常存活ꎬ这可能与植物的光修复有关ꎬ因此ꎬ
下降有关ꎮ 研究表明ꎬ在胁迫环境下ꎬ环式电子传 可在高温强光下对短命植物光损伤的核心原因ꎬ
递流可通过电子从 PS I 传递至 PQ 处建立高的类 如 VDE 活性和 PsbS 降解等ꎬ以及光修复机制等方
囊体膜质子梯度( ΔpH)ꎬ而这一梯度可促进调节 面进行进一步的研究ꎬ有助于更加全面了解短命
性能量耗散ꎬ提高 Y(NPQ)ꎬ保护 PS Ⅱ、PS I 免受 植物光合适应机理ꎮ