
Page 29 - 《广西植物》2024年第2期
P. 29
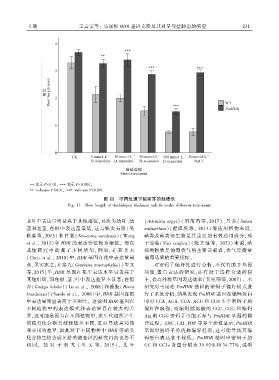
2 期 王嘉雯等: 马尾松 HDR 基因克隆及其对旱与盐胁迫的响应 2 3 1
∗∗表示 P<0.01ꎬ∗∗∗表示 P<0.001ꎮ
∗∗ indicates P<0.01ꎬ ∗∗∗ indicates P<0.001.
图 11 不同处理下拟南芥的胚根长
Fig. 11 Root length of Arabidopsis thaliana radicle under different treatments
老叶中表达量明显高于其他部位ꎬ其次为幼叶、幼 (Artemisia argyi) ( 刘 苗 苗 等ꎬ2017)、 丹 参 ( Salvia
茎和老茎ꎬ在根中表达量最低ꎬ这与铁皮石斛( 吴 miltiorrhiza)(程琪庆等ꎬ 2013) 等药用植物来说ꎬ
秋菊等ꎬ2015) 和甘薯( Dioscorea esculenta) ( Wang 萜类及萜类衍生物是其重要的有效药用成分ꎻ对
et al.ꎬ 2012) 中 HDR 的表达特征较为相似ꎮ 而在 于葡萄( Vitis vinifera) ( 陈天池等ꎬ 2023) 来说ꎬ萜
其他 研 究 中 出 现 了 不 同 结 果ꎬ 例 如ꎬ 在 萝 芙 木 烯类物质是葡萄香气的主要贡献者ꎬ香气是衡量
(Chen et al.ꎬ 2010)中ꎬHDR 基因在花中表达量最 葡萄品质的重要指标ꎮ
高ꎬ果实次之ꎻ在秦艽(Gentiana macrophylla) ( 岑文 对密码子偏好性进行分析ꎬ不仅有助于基因
等ꎬ2015)中ꎬHDR 基因在花中表达水平显著高于 功能、蛋白表达的研究ꎬ亦有助于选择合适的宿
其他组织ꎬ而在根、茎、叶中表达差异不显著ꎻ在银 主ꎬ提高外源基因表达效率( 吴宪明等ꎬ2007)ꎮ 本
杏(Ginkgo biloba) ( Lu et al.ꎬ 2008) 和橡胶( Hevea 研究对马尾松 PmHDR 基因的密码子偏好模式进
brasiliensis)(Sando et al.ꎬ 2008) 中ꎬHDR 基因在根 行了系统分析ꎬ结果发现 PmHDR 基因在编码蛋白
中表达量则显著高于茎和叶ꎮ 这说明 HDR 基因在 时对 CCA、AGA、GGA、ACU 和 CUU 5 个密码子的
不同植物中的表达模式和表达量存在较大的差 偏好性极强ꎬ而编码脯氨酸的 CCC、CCG 和编码
异ꎬ这可能是因为在不同植物中ꎬ次生代谢所产生 Arg 的 CGG 密码子可能不参与 PmHDR 基因的翻
的萜类化合物类型和数量不同ꎬ进而导致其功能 译过程ꎮ ENC、CAI、FOP 等多个参数显示ꎬPmHDR
和应用的差异ꎬ因此对于不同物种中 HDR 等萜类 基因对密码子的选择偏好性弱ꎬ这可能导致其编
化合物生物合成上游关键基因的研究目的也各不 码蛋白表达水平较低ꎮ PmHDR 基因中密码子的
相 同ꎮ 如 对 于 秦 艽 ( 岑 文 等ꎬ 2015 )、 艾 叶 GC 和 GC3s 含量分别为 39.92%和 34.77%ꎬ说明