
Page 27 - 《广西植物》2024年第2期
P. 27
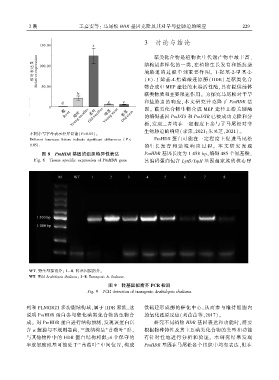
2 期 王嘉雯等: 马尾松 HDR 基因克隆及其对旱与盐胁迫的响应 2 2 9
3 讨论与结论
萜类化合物是植物次生代谢产物中最丰富、
结构最多样化的一类ꎬ在植物生长发育和抵抗逆
境胁 迫 的 过 程 中 到 重 要 作 用ꎮ 1 ̄羟 基 ̄2 ̄甲 基 ̄2 ̄
(E) ̄丁烯基 ̄4 ̄焦磷酸还原酶( HDR) 是萜类化合
物合成中 MEP 途径的末端活性酶ꎬ具有提供前体
萜类物质和主要限速作用ꎮ 为探究马尾松对干旱
和盐胁迫的响应ꎬ本文研究并克隆了 PmHDR 基
因ꎮ 萜类化合物生物合成 MEP 途径上游关键酶
的编码基因 PmDXS 和 PmDXR 已被成功克隆和分
析ꎬ发现二者均在一定程度上参与了马尾松对非
生物胁迫的响应(李荣ꎬ2021ꎻ朱灵芝ꎬ2021)ꎮ
不同小写字母表示差异显著(P<0.05)ꎮ
PmHDR 蛋白可能在一定程度上促进马尾松
Different lowercase letters indicate significant differences ( P <
0.05). 的生长 发 育 和 逆 境 响 应 过 程ꎮ 本 文 研 究 发 现
图 8 PmHDR 基因的组织特异性表达 PmHDR 基因长度为 1 458 bpꎬ编码 485 个氨基酸ꎬ
Fig. 8 Tissue specific expression of PmHDR gene 其编码蛋白包含 LytB / IspH 基因超家族的核心序
WT. 野生型拟南芥ꎻ 1~ 8. 转基因拟南芥ꎮ
WT. Wild Arabidopsis thalianaꎻ 1-8. Transgenic A. thaliana.
图 9 转基因拟南芥 PCR 检测
Fig. 9 PCR detection of transgenic Arabidopsis thaliana
列和 PLN02821 多功能结构域ꎬ属于 HDR 家族ꎬ这 铁硫建形成酶的催化中心ꎬ从而参与维持细胞内
说明 PmHDR 蛋白参与催化萜类化合物的生物合 的氧化还原反应(刘苗苗等ꎬ2017)ꎮ
成ꎮ 对 PmHDR 蛋白进行结构预测ꎬ发现该蛋白富 研究不同植物 HDR 基因表达和功能时ꎬ需要
含 α 螺旋与不规则卷曲ꎬ三级结构呈“苜蓿叶” 形ꎬ 根据物种特性及其主要萜类化合物的类型和功能
与其他物种中的 HDR 蛋白结构相似ꎻ4 个保守的 有针对性地进行分析和验证ꎮ 本研究结果发现
半胱氨酸残基可能处于“ 苜蓿叶” 中间位置ꎬ构成 PmHDR 基因在马尾松各个组织中均有表达ꎬ但在