
Page 76 - 《广西植物》2024年第2期
P. 76
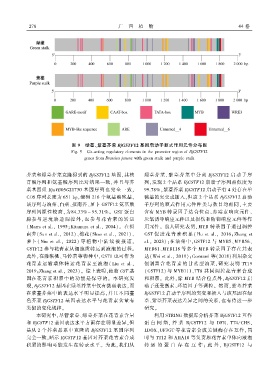
2 7 8 广 西 植 物 44 卷
图 9 绿薹、紫薹芥菜 BjGSTF12 基因启动子顺式作用元件分布图
Fig. 9 Cis ̄acting regulatory elements in the promoter region of BjGSTF12
genes from Brassica juncea with green stalk and purple stalk
芥菜和绿薹芥菜克隆得到的 BjGSTF12 基因ꎬ其核 绿薹芥菜、紫 薹 芥 菜 中 分 离 BjGSTF12 启 动 子 序
苷酸序列和氨基酸序列比对结果一致ꎬ并且与芥 列ꎬ发现 2 个品系 BjGSTF12 启动子序列相似度为
菜基因组 BjuVB05G21730 基因序列也完全一致ꎬ 99.78%ꎬ紫薹芥菜 BjGSTF12 启动子有 4 处存在单
CDS 序列长度为 651 bpꎬ编码 216 个氨基酸残基ꎬ 碱基的突变或插入ꎬ但这 2 个品系 BjGSTF12 启动
该序列与油菜、白菜、拟南芥、萝卜 GSTF12 氨基酸 子序列的顺式作用元件种类与数目均相同ꎬ主要
序列同源性较高ꎬ为84.33% ~ 95.31%ꎮ GST 蛋白 含有 MYB 转录因子结合位点、赤霉素响应元件、
除参与 逆 境 胁 迫 调 控 外ꎬ 也 参 与 花 青 素 的 转 运 厌氧诱导响应元件以及损伤和防御响应元件等作
(Marrs et al.ꎬ 1995ꎻKitamura et al.ꎬ 2004)ꎮ 在拟 用元件ꎮ 前人研究表明ꎬMYB 转录因子通过调控
南芥(Sun et al.ꎬ 2012)、棉花(Shao et al.ꎬ 2021)、 GST 促 进 花 青 素 积 累 ( Hu et al.ꎬ 2016ꎻ Zhang et
萝卜 ( Niu et al.ꎬ 2022) 等 植 物 中 陆 续 被 报 道ꎬ al.ꎬ 2023)ꎻ在油菜中ꎬGSTF12 与 MYB5、MYB56、
GSTF12 参与花青素从细胞质转运到液泡的过程ꎮ MYB61、MYB118 等多个 MYB 转录因子存在共表
此外ꎬ在猕猴桃、马铃薯等物种中ꎬGST1 也可作为 达(Wei et al.ꎬ 2019)ꎻGoswami 等(2018)利用杂交
花青素运输载体转 运 花 青 素 至 液 泡 ( Liu et al.ꎬ 创制富 含 花 青 素 的 甘 蓝 型 油 菜ꎬ研 究 表 明 TT19
2019ꎻZhang et al.ꎬ 2023)ꎮ 综上表明ꎬ植物 GST 基 (GSTF12) 与 MYB111、TT8 共同调控花青素合成
因在花青素积累中的功能是保守的ꎮ 本研究发 和积累ꎮ 此外ꎬ除 MYB 结合位点外ꎬBjGSTF12 启
现ꎬBjGSTF12 基因在绿薹芥菜中仅有微弱表达ꎬ而 动子还受激素、环境因子等调控ꎮ 然而ꎬ紫薹芥菜
在紫薹芥菜中的表达水平明显提高ꎬ并且不同薹 BjGSTF12 启动子序列的突变和插入与该基因在绿
色芥菜 BjGSTF12 基因表达水平与花青素含量有 薹、紫薹芥菜表达差异之间的关系ꎬ也有待进一步
类似的变化规律ꎮ 研究ꎮ
本研究中ꎬ尽管紫薹、绿薹芥菜在花青素含量 利用 STRING 数据库分析芥菜 BjGSTF12 互作
和 BjGSTF12 基因表达水平方面存在明显差异ꎬ但 蛋 白 网 络ꎬ 芥 菜 BjGSTF12 与 DFR、 TT4 / CHS、
是从 2 个芥菜品系中克隆的 BjGSTF12 基因序列 LDOX、UF3GT 等花青素合成关键酶存在互作ꎬ同
完全一致ꎬ暗示 BjGSTF12 基因对芥菜花青素合成 时与 TT12 和 AHA10 等负责原花青素单体向液泡
积累的影响可能发生在转录水平ꎮ 为此ꎬ我们从 转 运 的 蛋 白 存 在 互 作ꎻ 此 外ꎬ BjGSTF12 与