
Page 192 - 《广西植物》2025年第10期
P. 192
1 9 2 2 广 西 植 物 45 卷
化应激可能是由羟基自由基所主导ꎮ 本研究检测
了与氧化应激相关的 11 个指标ꎬ其中冻存后抑制
羟基自由基的能力较对照显著下降ꎮ 此外ꎬSOD、
GSH、APX 等指标含量或活性也较对照发生了显
著变化ꎬ但 MDA 含量没有显著变化ꎮ GSH 是植物
内重要的小分子抗氧化物质ꎮ GSH 代谢可有效清
除植物过量的过氧化物ꎬ在植物的信号传导和逆
境胁迫响应中具有重要作用( Matés et al.ꎬ 2002)ꎮ
这表明抗氧化酶和抗氧化物质参与了冷冻保存细
胞保护机制ꎮ
转录组学被广泛应用于动植物发育调控、免
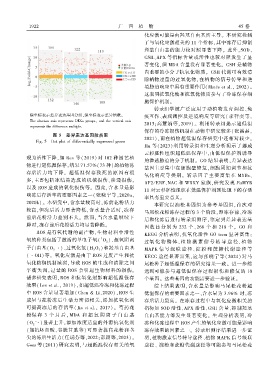
横坐标表示差异表达基因分组ꎬ纵坐标表示差异倍数ꎮ
疫互作、表观调控及逆境响应等研究( 李智奕等ꎬ
The abscissa axis represents DEGs groupsꎬ and the vertical axis
2013ꎻ高慧娟等ꎬ2019)ꎮ 利用转录组揭示超低温
represents the difference multiple.
保存的冷冻损伤机制在动物中研究较多( 张晶晶ꎬ
图 5 差异表达基因散点图
2021)ꎬ而在植物超低温保存研究中还相对较少ꎮ
Fig. 5 Dot plot of differentially expressed genes
Hu 等(2023)利用转录组和生理分析揭示了挪威
云杉胚性组织超低温保存中ꎬ由低温保护剂诱导
现为活性下降ꎬ如 Ren 等(2019) 对 102 种园艺植
的渗透胁迫的分子机制ꎮ GO 结果表明ꎬ差异表达
物进行超低温保存ꎬ结果 71.57%(73 种)的植物冻
基因主要集中在细胞壁修复ꎬ细胞周期调节和抗
存后活力均下降ꎮ 超低温保存致死的原因有很
氧化响应等类别ꎮ 转录因子主要富集在 MYBs、
多ꎬ主要包括冰结晶造成的机械损伤、渗透损伤ꎬ
AP2 / ERF、NAC 和 WRKY 家族ꎬ研究发现 PaMYB
以及 ROS 造成的氧化损伤等ꎮ 因此ꎬ含水量是影
11 对云杉胚性组织在低温保护剂预处理下的存活
响冻后存活率的重要因素之一( 张晓宁等ꎬ2020aꎬ
率具有重要意义ꎮ
2020b)ꎮ 本研究中ꎬ含水量较高时ꎬ冻前花粉活力
本研究以油松基因组为参考基因组ꎬ首次对
较高ꎬ但冻后活力显著降低ꎬ含水量合适时ꎬ冻存 马尾松花粉冻存过程的 3 个阶段ꎬ即冻存前、冷冻
前后花粉活力差别不大ꎮ 然而ꎬ当含水量继续下 后和化冻后进行转录组测序ꎬ鉴定到差异表达基
降时ꎬ冻存前后花粉活力均显著降低ꎮ
因数 目 分 别 为 232 个、268 个 和 218 个ꎮ GO 和
ROS 是有氧代谢的副产物ꎬ生物材料中活性
KEGG 分析表明ꎬ抗氧化活性 GO term 显著富集ꎻ
氧的种类包括了激活的单电子氧( O )、超氧阴离
1
2 过氧 化 物 酶 体、 植 物 激 素 信 号 转 导 途 径、 植 物
子自由基(O )、过氧化氢(H O ) 和羟基自由基 MAPK 信 号 级 联 途 径、 淀 粉 和 蔗 糖 代 谢 途 径 等
-
2 2 2
(OH)等ꎮ 氧化应激是由于 ROS 过度产生和抗 KEGG 途径显著富集ꎬ这与张晓宁等(2024) 对马
氧化防御机制减弱ꎬ导致 ROS 的生成和清除之间 尾松种子超低温保存的研究结果一致ꎮ 进一步筛
平衡失调ꎬ过量的 ROS 会引起生物材料的损伤ꎮ 选到可能参与超低温保存过程损伤和修复的 10
诸多研究表明ꎬROS 和抗氧化剂影响超低温保存 个基因ꎮ 这些基因的功能需要进一步验证ꎮ
效果(Len et al.ꎬ 2019)ꎬ如超低温冷冻和化冻过程 综上结果表明ꎬ含水量是影响马尾松花粉超
中 ROS 含量显著增加( Chen & Liꎬ2020)ꎬROS 生 低温保存的重要因素之一ꎬ含水量为 3.96% 时ꎬ冻
成量与花粉冻后生活力密切相关ꎬ添加抗氧化剂 存后活力最高ꎮ 在冻存过程中与氧化应激相关的
可提高冻后的存活率( Jia et al.ꎬ 2017)ꎮ 芍药花 指标如 SOD 活性、APX 活性、GSH 含量、抑制羟基
粉保 存 3 个 月 后ꎬ MDA 和 超 氧 阴 离 子 自 由 基 自由基能力等发生显著变化ꎮ 生理分析表明ꎬ冷
(O )显著上升ꎬ添加浓度适宜的外源抗氧化剂 冻和化冻过程中 ROS 产生的氧化应激可能是影响
-
2
(如抗坏血酸、谷胱甘肽等) 可显著提高花粉和茎 冻存效果的因素之一ꎮ 转录组测序结果进一步证
尖的冻后生活力(任瑞芬等ꎬ2022ꎻ彭颖等ꎬ2023)ꎮ 明ꎬ植物激素信号转导途径、植物 MAPK 信号级联
Goes 等(2011) 研究表明ꎬ与超低温保存相关的氧 途径、 淀粉和蔗糖代谢途径等可能参与马尾松花