
Page 50 - 《广西植物》2025年第11期
P. 50
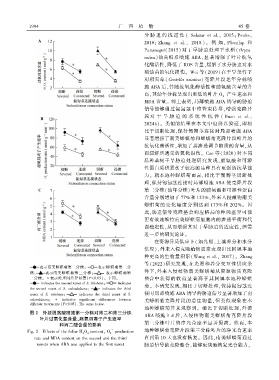
1 9 9 4 广 西 植 物 45 卷
分 胁 迫 的 抗 逆 性 ( Salazar et al.ꎬ 2015ꎻ Peukeꎬ
2016ꎻ Zhang et al.ꎬ 2018 )ꎮ 例 如ꎬ Ploenlap 和
Pattanagul(2015) 对干旱胁迫处理下水稻( Oryza
sativa)幼苗根系喷施 ABAꎬ显著增强了叶片抗氧
化酶活性ꎬ降低了 ROS 含量ꎬ缓解了水分胁迫对水
稻幼苗的氧化损伤ꎮ Wei 等(2019) 在干旱条件下
对积雪草( Centella asiatica) 克隆片段老年分株喷
施 ABA 后ꎬ伴随抗氧化酶活性和脯氨酸含量的升
高ꎬ其幼年分株呈现出更低的叶片 O 产生速率和
-
2
MDA 含量ꎮ 综上表明ꎬ局部喷施 ABA 诱导的胁迫
信号能够通过匍匐茎中维管束传导ꎬ增强克隆片
段 对 干 旱 胁 迫 的 系 统 性 抗 性 ( Duan et al.ꎬ
2024b)ꎮ 类似的结果在本文中也得以验证ꎬ即相
比于切断处理ꎬ保持匍匐茎连接时局部喷施 ABA
显著增强了南美蟛蜞菊和蟛蜞菊克隆片段叶片的
抗氧化酶活性ꎬ增加了其渗透调节物质的含量ꎬ从
而缓解所遭受的氧化损伤ꎮ Cao 等(2020) 对不同
品种桑树干旱胁迫处理研究发现ꎬ脯氨酸和可溶
性蛋白质积累水平较高的品种具有更强的抗旱能
力ꎮ 就本地种蟛蜞菊而言ꎬ相比于匍匐茎切断处
理ꎬ保持匍匐茎连接时局部喷施 ABA 使克隆片段
第三分株(幼年分株)叶片的脯氨酸和可溶性蛋白
含量分别增加了 77%和 123%ꎬ外来入侵植物南美
蟛蜞菊的变化幅度分别达到 173% 和 202%ꎮ 因
此ꎬ胁迫信号克隆整合响应格局的种间差异可能
更有效地维持南美蟛蜞菊细胞内的渗透平衡和代
谢稳定性ꎬ从而增强其对干旱胁迫的适应性ꎬ但需
进一步的研究论证ꎮ
在资源异质供应下(如光照、土壤养分和水分
供应)ꎬ外来入侵克隆植物通常表现出比同属本地
种更高的生物量积累( Wang et al.ꎬ 2017)ꎮ Zhang
等(2022)研究发现ꎬ在光照和养分交互性供应条
表示南美蟛蜞菊第二分株ꎻ 表示蟛蜞菊第二分
株ꎻ 表示南美蟛蜞菊第三分株ꎻ 表示蟛蜞菊第 件下ꎬ外来入侵植物南美蟛蜞菊从资源物质克隆
三分株ꎮ ∗表示处理间差异显著(P<0.05)ꎮ 下同ꎮ 整合中获得的收益显著高于其同属本地种蟛蜞
indicates the second ramet of S. trilobataꎻ indicates
菊ꎮ 本研究发现ꎬ相比于切断处理ꎬ保持匍匐茎连
the second ramet of S. calendulaceaꎻ indicates the third
ramet of S. trilobataꎻ indicates the third ramet of S. 接时局部喷施 ABA 诱导的胁迫信号显著增加了南
calendulaceaꎻ ∗ indicates significant differences between 美蟛蜞菊克隆片段的总生物量ꎬ但类似现象在本
different treatments (P<0.05). The same below.
地种蟛蜞菊并未观察到ꎮ 相比于切断处理ꎬ外源
图 2 外源脱落酸喷施第一分株对第二和第三分株 ABA 喷施 3 d 后ꎬ入侵植物南美蟛蜞菊克隆片段
叶片过氧化氢含量、超氧阴离子产生速率
和丙二醛含量的影响 第三分株叶片的净光合速率显著提高ꎮ 然而ꎬ本
- 地种蟛蜞菊克隆片段第三分株叶片的净光合速率
Fig. 2 Effects of the foliar H O contentꎬ O production
2 2 2
rate and MDA content on the second and the third 直到第 10 天也没有恢复ꎮ 因此ꎬ南美蟛蜞菊通过
ramets when ABA was applied to the first ramet 胁迫信号的克隆整合ꎬ能够更快地恢复光合能力ꎬ