
Page 149 - 《广西植物》2025年第12期
P. 149
12 期 刘依婷等: 低温胁迫下兴安白芷与川白芷种子转录组特征及激素调控基因的比对研究 2 2 9 1
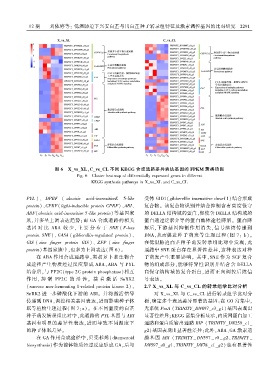
图 6 X_vs_XL、C_vs_CL 不同 KEGG 合成通路差异表达基因的 FPKM 聚类热图
Fig. 6 Cluster heatmap of differentially expressed genes in different
KEGG synthesis pathways in X_vs_XL and C_vs_CL
PYL )、 DPBF ( abscisic acid ̄insensitiveE 5 ̄like 受体 GID1(gibberellin insensitive dwarf 1)结合形成
protein)、CPRF( light ̄inducible protein CPRF)、ABI、 复合物ꎮ 该复合物识别并结合抑制含有高度保守
ABF(abscisic acid ̄insensitive 5 ̄like protein) 等基因家 的 DELLA 结构域的蛋白ꎬ促使含 DELLA 结构域的
族ꎬ并多呈上调表达趋势ꎻ而 GA 合成通路的相关 蛋白通过泛素介导的蛋白酶体途径降解ꎮ 蛋白降
基因 对 比 ABA 较 少ꎬ 主 要 分 布 于 SNE ( F ̄box 解后ꎬ下游基因抑制作用消失ꎬ信号继续传递到
protein SNE)、 GASA ( gibberellin ̄regulated protein)、 DNAꎬ从而促进种子萌发等生理过程( 图 7: b)ꎮ
GIS ( zinc finger protein GIS )、 ZFP ( zinc finger 在低温胁迫白芷种子萌发转录组比对中发现ꎬ此
protein)基因家族中ꎬ也多为上调表达(图 6)ꎮ 通路中 SNE 蛋白存在显著性差异ꎬ这种表达对种
在 ABA 作用合成通路中ꎬ类胡萝卜素生物合 子萌发产生重要影响ꎮ 其中ꎬSNE 作为 SCF 复合
成途径产生物质经过反应形成 ABAꎬABA 与 PYL 物的组成部分ꎬ能够特异性识别并结合含 DELLA
结合后ꎬ与 PP2C(type 2C protein phosphatase)相互 高保守结构域的复合蛋白ꎬ进而正向调控后续信
作用ꎬ 抑 制 PP2C 的 活 性ꎮ 接 着 激 活 SnRK2 号表达ꎮ
(sucrose non ̄fermenting 1 ̄related protein kinase 2)ꎬ 2.7 X_vs_XL 与 C_vs_CL 的转录组学比对分析
SnRK2 进一步磷酸化下游的 ABIꎮ 并将激活信号 对 X_vs_XL 与 C_vs_CL 进行转录组学比对分
传递到 DNAꎬ调控相关基因表达ꎬ进而影响种子休 析ꎬ确定多个表达差异显著的基因:在 GO 富集中ꎬ
眠等植物生理过程( 图 7:a)ꎮ 在不同温度的白芷 光系统 PsaA (TRINITY_DN897_c3_g1)基因表现出
种子萌发转录组比对中ꎬ此通路的 PYL 基因与 ABI 显著性差异ꎻKEGG 富集分析显示ꎬ内质网蛋白加工
基因有明显的差异性表达ꎬ进而导致不同温度下 通路和蛋白质输出通路 BIP (TRINITY_DN259_c1_
的种子休眠差异ꎮ g2)基因表现出显著性差异ꎻ此外ꎬABA、GA 激素通
在 GA 作用合成途径中ꎬ贝壳杉烯( diterpenoid 路基 因 ABI ( TRINITY _ DN597 _ c0 _ g2、 TRINITY _
biosynthesis)作为前体物质经过反应形成 GAꎬ后与 DN597_c0_g1、TRINITY_DN78_c1_g2) 也有显著性