
Page 183 - 《广西植物》2026年第1期
P. 183
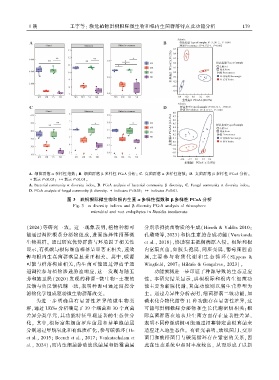
1 期 王宇等: 濒危植物珙桐根际微生物和根内生菌群落特点及功能分析 1 7 9
A. 细菌群落 α 多样性指数ꎻ B. 细菌群落 β 多样性 PCoA 分析ꎻ C. 真菌群落 α 多样性指数ꎻ D. 真菌群落 β 多样性 PCoA 分析ꎮ
∗表示 P<0.05ꎻ ∗∗表示 P<0.01ꎮ
A. Bacterial community α diversity indexꎻ B. PCoA analysis of bacterial community β diversityꎻ C. Fungal community α diversity indexꎻ
D. PCoA analysis of fungal community β diversity. ∗ indicates P<0.05ꎻ ∗∗ indicates P<0.01.
图 3 珙桐根际微生物和根内生菌 α 多样性指数和 β 多样性 PCoA 分析
Fig. 3 α diversity indices and β diversity PCoA analysis of rhizosphere
microbial and root endophytes in Davidia involucrata
(2024)等研究一致ꎮ 这一现象表明ꎬ植物种源可 分别承担抗菌物质的生成( Hirsch & Valdés 2010ꎻ
能通过调控根系分泌物组成ꎬ进而选择性招募微 孔璐琦等ꎬ2023) 和抗生素的合成功能( Vurukonda
生物类群ꎮ 通过研究优势群落与环境因子相关性 et al.ꎬ 2018)ꎬ协助宿主抵御病原入侵ꎮ 根际和根
显示ꎬ有机碳与根际细菌群落呈显著正相关ꎬ速效 内富集真菌ꎬ如胶头孢属、间座壳属、葡萄座腔菌
钾与根内生真菌群落呈显著正相关ꎮ 其中ꎬ碳源 属ꎬ主要 参 与 物 质 代 谢 和 生 态 循 环 ( Slippers &
可能与群落构建相关ꎬ内生菌可能通过钾离子通 Wingfieldꎬ 2007ꎻ Hilário & Gonçalvesꎬ 2023)ꎮ
道调控参与植物渗透胁迫响应ꎬ这一发现与陆玉 功能预测进一步印证了种源导致的生态适应
芳和施卫民(2020) 发现的种源-微生物 -土壤的 性ꎮ 本研究结果显示ꎬ珙桐根际和根内生细菌功
反馈与负反馈机制一致ꎬ表明种源可通过调控分 能主要为新陈代谢ꎬ真菌功能则以腐生营养型为
泌物化学组成驱动微生物群落改变ꎮ 主ꎮ 通过差异性分析表明ꎬ细菌群落二级功能ꎬ如
为进一步明确具有显著 性 差 异 的 微 生 物 类 碳水化合物代谢等 11 种功能存在显著性差异ꎬ这
群ꎬ通过 LEfSe 分析确定了 39 个细菌和 30 个真菌 可能与珙桐根际分泌物和生长代谢密切相关ꎻ根
差异分类单元ꎬ其功能特征呈现显著的生态位分 际真菌群落在地衣共生菌方面存在显著性差异ꎬ
化ꎮ 其中ꎬ根际富集细菌罗库菌属和芽单胞菌属 表明不同种源珙桐可能通过招募特定菌根真菌来
分别通过甲烷氧化和有机质矿化ꎬ参与碳循环( He 适应迁入地生态位ꎮ 有研究表明ꎬ放线菌门、变形
et al.ꎬ 2015ꎻ Becraft et al.ꎬ 2017ꎻ Venkatachalam et 菌门和酸杆菌门与碳氮循环存在紧密的关系ꎬ因
al.ꎬ 2024)ꎻ而内生细菌游动放线菌属和链霉菌属 此在生态系统中相对丰度较高ꎬ 从而形成了以新