
Page 76 - 《广西植物》2026年第1期
P. 76
7 2 广 西 植 物 46 卷
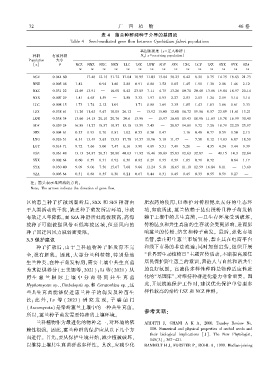
表 4 蕙兰种群间种子介导的基因流
Table 4 Seed ̄mediated gene flow between Cymbidium faberi populations
基因流强度 [x = 迁入种群]
种群 有效种群 N m [x = receiving population]
Population 大小
[x] θ NCZ NNZ NXC NXX LLC LSX LXW SDF SFS LXG LGY LSZ SXX SYX SZA
→x →x →x →x →x →x →x →x →x →x →x →x →x →x →x
NCZ 0.044 60 — 17.48 12.15 31.72 17.04 18.95 11.03 13.84 30.23 8.42 8.30 8.75 14.75 16.63 21.73
NNZ 0.005 08 1.42 — 0.94 1.60 2.88 0.91 0.88 1.52 1.85 1.45 1.91 1.30 2.08 1.46 2.12
NXC 0.031 22 12.69 13.91 — 16.00 8.42 23.05 7.11 4.75 13.26 10.71 20.48 13.66 19.84 18.97 20.14
NXX 0.007 29 1.41 4.65 1.59 — 2.58 3.32 1.97 0.93 2.27 2.53 2.03 1.24 2.19 3.14 3.14
LLC 0.008 15 1.73 1.74 2.12 1.01 — 1.71 0.88 1.69 3.35 1.05 1.43 1.63 3.66 0.61 3.33
LSX 0.038 61 11.26 11.63 9.67 18.05 26.12 — 13.92 15.08 12.68 16.72 19.56 8.57 23.09 11.61 13.21
LXW 0.038 34 13.66 14.15 20.10 20.76 20.6 13.96 — 15.97 16.68 15.43 18.96 11.63 15.78 16.99 30.40
SDF 0.039 24 16.88 11.27 18.57 10.57 13.18 13.59 7.45 — 28.07 14.81 9.72 7.26 18.79 22.25 25.97
SFS 0.004 61 0.55 0.93 0.70 0.81 1.02 0.55 0.50 0.47 — 1.16 0.48 0.77 0.59 0.50 2.13
LXG 0.026 51 6.34 13.49 5.65 15.93 17.78 14.57 10.96 5.18 11.47 — 7.58 8.12 14.03 6.87 18.50
LGY 0.014 71 9.72 7.66 5.06 7.47 6.16 3.98 4.69 5.51 7.49 5.26 — 4.35 4.24 3.44 9.39
LSZ 0.061 48 13.13 24.87 20.51 20.86 40.03 11.92 16.46 38.69 25.03 12.63 32.87 — 40.15 14.3 22.64
SXX 0.002 64 0.60 0.59 0.31 0.92 0.50 0.82 0.29 0.95 0.59 1.05 0.71 0.72 — 0.64 1.17
SYX 0.030 40 9.09 9.06 7.56 15.67 7.68 9.66 11.24 5.18 18.65 11.10 12.59 16.84 8.11 — 13.60
SZA 0.003 64 0.31 0.68 0.35 0.30 0.21 0.67 0.44 0.31 0.45 0.45 0.33 0.55 0.59 0.27 —
注: 箭头表示基因流的方向ꎮ
Note: The arrows indicate the direction of gene flow.
区的蕙兰种子扩散到秦岭后ꎬSXX 和 SFS 种群由 肥农药的使用ꎬ以维护对传粉昆虫友好的生态环
于人类活动的干扰ꎬ缺乏种子萌发所需环境ꎬ导致 境ꎬ如前所述ꎬ蕙兰依赖于昆虫授粉且种子萌发依
有效迁入率降低ꎬ而 SZA 种群所处海拔较高ꎬ高海 赖于土壤中的共生真菌ꎬ一旦生存环境受到破坏ꎬ
拔种子可能被强风带至低海拔区域ꎬ但逆风向的 传粉昆虫和共生真菌的生存就会受到影响ꎬ进而影
种子回迁因风力减弱而受限ꎮ 响蕙兰的传粉、结实和种子萌发ꎮ 最后ꎬ强化市场
3.3 保护建议 监管ꎬ禁止野生蕙兰市场贸易ꎬ禁止其在电商平台
种子扩散后ꎬ由于兰科植物种子胚发育不完 和线下市场的非法流通ꎻ同时加强宣传ꎬ组织开展
全ꎬ没有胚乳ꎮ 因此ꎬ大部分兰科植物ꎬ特别是地 “世界野生动植物日”主题宣传活动ꎬ不断提高原住
生兰种类ꎬ在种子萌发初期ꎬ需要土壤中共生真菌 居民保护野生蕙兰的意识ꎬ营造人与自然和谐共生
为其提供养分( 王美娜等ꎬ2021)ꎻLi 等(2021) 从 的良好氛围ꎮ 高遗传多样性种群是物种适应性进
野 生 蕙 兰 根 际 土 壤 中 分 离 得 到 共 生 真 菌 化的“基因库”ꎬ对维持种群进化潜力非常重要ꎮ 因
Hyphomycete sp.、Umbelopsis sp.和 Ceratorhiza sp.ꎬ这 此ꎬ开展就地保护工作时ꎬ建议优先保护单倍型多
些共生真菌能够促进蕙兰种子的萌发及种苗生 样性较高的秦岭 LSZ 和 NCZ 种群ꎮ
长ꎻ 此 外ꎬ Lv 等 ( 2023 ) 研 究 发 现ꎬ 子 囊 菌 门
(Ascomycota)是秦岭蕙兰土壤中的一种共生真菌ꎮ
参考文献:
所以ꎬ蕙兰种子萌发需要特殊的土壤环境ꎮ
兰科植物作为最进化的物种之一ꎬ对环境的依 ARDITTI Jꎬ GHANI A K Aꎬ 2000. Tansley Review No.
赖性很强ꎮ 因此ꎬ蕙兰种群的保护应从以下几个方 110. Numerical and physical properties of orchid seeds and
their biological implications [ J]. The New Phytologistꎬ
面进行ꎮ 首先ꎬ要从保护生境开始ꎬ减少植被破坏ꎬ
145(3): 367-421.
以维持土壤共生真菌群落多样性ꎮ 其次ꎬ应减少化 BANDELT H Jꎬ FORSTER Pꎬ ROHL Aꎬ 1999. Median ̄joining