
Page 140 - 《广西植物》2026年第5期
P. 140
8 7 2 广 西 植 物 46 卷
本研究对氮素代谢途径差异表达基因的表达
模式分析表明ꎬ有 88 个差异表达基因参与了氮素
代谢途径的 11 个主要环节ꎮ 这些基因主要涉及
硝酸转运蛋白、硝酸还原酶、亚硝酸还原酶、谷氨
酸合成酶等关键酶类ꎮ 在对正常氮条件下的小麦
进行转录组分析时ꎬ也鉴定到这些参与氮代谢途
径的关键酶(Livak & Schmittgenꎬ 2001ꎻ Yin et al.ꎬ
2019)ꎮ 部分氮代谢途径相关基因的表达量随着
铵离子浓度的增加而发生显著变化ꎬ这表明铵离
子浓 度 对 氮 代 谢 途 径 的 调 控 具 有 重 要 作 用
(Goodstein et al.ꎬ 2012ꎻWang et al.ꎬ 2019)ꎮ 这种
调控机制可能与植物在不同氮素形态下ꎬ通过精
细调节关键酶的活性和表达水平来优化氮素的同
化和利用效率有关ꎬ从而确保植物在不同氮素供
应条件下的生长和发育(Yang et al.ꎬ 2025)ꎮ
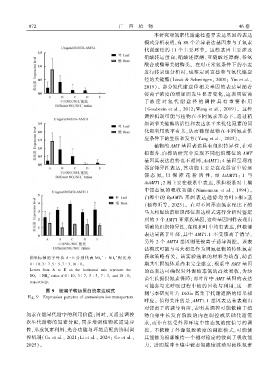
植物的 AMT 基因表达具有组织特异性ꎬ在对
拟南芥、白桦的研究中发现不同组织部位的 AMT
基因其表达趋势也不相同ꎬAtAMT1ꎻ4 基因呈现花
器官特异性表达ꎬ其功能主要是在花器官中转运
铵 态 氮ꎬ 以 保 障 花 粉 活 性ꎬ 而 AtAMT1ꎻ 1 与
AtAMT1ꎻ2 则主要在根系中表达ꎬ承担根系对土壤
中铵态氮的吸收功能( Ninnemann et al.ꎬ 1994)ꎮ
白桦中的 BpAMTs 基 因 表 达 趋 势 均 为 叶 > 根 > 茎
(杨海昕等ꎬ 2023)ꎮ 在对不同形态氮素配比下的
马大相思幼苗组织部位表达模式进行分析时鉴定
到的 3 个 AMT1 亚家族基因ꎬ这些基因同样表现出
明确的组织特异性ꎬ在根和叶中均有表达ꎬ但根部
表达量高于叶部ꎬ其中 AMT1.1 不受铵离子诱导ꎬ
另外 2 个 AMTA 基因则受铵离子诱导调控ꎮ 该表
达模式可能与马大相思作为固氮植物的特殊氮素
- + 获取策略有关ꎮ 该实验选取的材料为幼苗ꎬ幼苗
横坐标 轴 的 字 母 从 A - E 分 别 代 表 NO 3 ∶ NH 4 配 比 为
0 ∶ 10、3 ∶ 7、5 ∶ 5、7 ∶ 3、10 ∶ 0ꎮ 期共生固氮体系尚未完全建立ꎬ根系中 AMT 基因
Letters from A to E on the horizontal axis represent the 的高表达可确保对外源铵态氮的高效吸收ꎬ为幼
- +
NO 3 ∶ NH 4 ratios of 0 ∶ 10ꎬ 3 ∶ 7ꎬ 5 ∶ 5ꎬ 7 ∶ 3ꎬ and 10 ∶ 0ꎬ
苗生长提供氮素保障ꎻ而叶片中 AMT 基因的表达
respectively.
可能参与光呼吸过程中铵的回收与利用ꎬ这一推
图 9 铵离子转运蛋白的表达模式
测与本研究叶片 DEGs 富集于代谢通路的结果相
Fig. 9 Expression patterns of ammonium ion transporters
呼应ꎮ 值得关注的是ꎬAMT1.1 基因表达未表现出
对铵离子的诱导响应ꎬ表明其调控可能依赖于植
氮素在能量代谢中的利用价值ꎻ同时ꎬ又通过调控 物自身生长发育阶段的内在调控或基础代谢需
次生代谢物的氮素分配ꎬ同步增强植物抗逆适应 求ꎬ而非直接受外界环境中铵态氮浓度信号的调
性ꎬ形成氮素利用、光合功能与环境适配的协同调 控ꎮ 不依赖于外源氮源浓度的调控模式ꎬ可能使
控机制(Gu et al.ꎬ 2021ꎻLi et al.ꎬ 2024ꎻ Ge et al.ꎬ 其能够为根部维持一个相对稳定的铵离子吸收能
2025)ꎮ 力ꎬ 进而缓冲土壤中铵态氮浓度波动对植株氮素