
Page 168 - 《广西植物》2026年第5期
P. 168
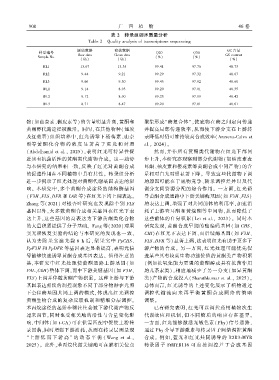
9 0 0 广 西 植 物 46 卷
表 2 转录组测序质量分析
Table 2 Quality analysis of transcriptome sequencing
原始数据 有效数据 GC 含量
样品编号 Q20 Q30
Raw data Clean data GC content
Sample No. (%) (%)
(Gb) (Gb) (%)
RL1 13.67 13.35 99.41 97.76 48.73
RL2 9.44 9.21 99.29 97.32 48.67
RL3 9.66 9.30 99.45 97.92 48.68
WL1 9.14 8.93 99.20 97.01 48.35
WL2 9.12 8.90 99.25 97.19 48.42
WL3 8.71 8.47 99.20 97.01 48.61
物(如山奈素、槲皮素等)的含量明显升高ꎬ黄酮和 聚集形成“ 酶复合体”ꎬ使底物在酶之间定向传递
黄酮醇代谢途径被激活ꎮ 同时ꎬ在其他物种( 如埃 并提高局部传递效率ꎬ从而使下游分支在上游转
及红菊薯) 组织培养中ꎬ红光诱导下杨梅素、山奈 录降低时仍可维持较高合成效率(Aravena ̄Calvo et
酚等 黄 酮 化 合 物 的 浓 度 显 著 高 于 蓝 光 和 对 照 al.ꎬ 2024)ꎮ
(Abdelhamid et al.ꎬ 2025)ꎬ表明红光可特异性促 然而ꎬ并非所有黄酮类代谢物在红光下都同
进具有抗菌活性的黄酮类代谢物合成ꎮ 这一趋势 步上升ꎬ本研究亦观察到部分代谢物( 如柚皮素查
与本研究的结果相一致ꎬ反映了红光对黄酮合成 耳酮、柚皮素和樱花素等黄酮合成中间产物) 的含
的促进作用在不同植物中具有共性ꎮ 转录组分析 量相对白光对照显著下降ꎮ 导致这些代谢物下调
进一步揭示了红光处理对黄酮代谢基因表达的影 的原因可能在于底物竞争、转录调控差异以及代
响ꎮ 本研究中ꎬ多个黄酮合成途径的结构酶基因 谢分支间资源分配的综合作用ꎮ 一方面ꎬ红光诱
(F3H、FLS、DFR 和 LAR 等)在红光下均上调表达ꎮ 导黄酮合成通路中下游关键酶基因(如 F3H、FLS)
Zhang 等(2021) 对银杏叶研究也发现除个别 FLS 的表达上调ꎬ增强了对共同前体的利用率ꎬ加速消
基因以外ꎬ大多数黄酮合成相关基因在红光下表 耗了上游查耳酮和黄烷酮等中间物ꎬ从而降低了
达上升ꎬ这些基因的高表达为下游黄酮类化合物 这些前体的自身积累( Lei et al.ꎬ 2023)ꎮ 同时本
的大量积累提供了分子基础ꎮ Fang 等(2020)对果 研究发现ꎬ黄酮合成早期的酶编码基因( 如 CHS、
实光照恢复实验的结论与本研究的发现也一致ꎬ CHI)在红光下表达下调ꎬ而后续酶基因( 如 F3H、
认为去 除 果 实 遮 光 袋 6 h 后ꎬ 梨 果 实 中 PpCHS、 FLS、DFR 等)显著上调ꎬ这表明红光有助于更多下
PpF3H 和 PpDFR 等基因表达显著提高ꎬ表明光信 游产物的合成ꎮ 另一方面ꎬ红光处理可能优先促
号能够快速诱导黄酮合成基因表达ꎮ 值得注意的 进某些具有较高生物功能价值的黄酮类产物积累
是ꎬ本研究中红光处理使黄酮通路上游基因( 如 (例如抗氧化能力更强的黄酮醇或具有抗菌作用
PAL、C4H)整体下调ꎬ而中下游关键基因( 如 F3H、 的儿茶素类)ꎬ相应地减少了另一分支( 如异黄酮
FLS)上调并伴随黄酮产物积累ꎮ 这种上游与下游 类)产物的合成投入( Sharathkumar et al.ꎬ 2025)ꎮ
基因表达相反的调控现象不同于部分物种在光照 总体而言ꎬ红光诱导的上述变化展示了植物通过
下全程酶基因共同上调的模式ꎬ体现出红光调控 调控 代 谢 流 向 来 再 平 衡 黄 酮 合 成 网 络 的 策 略
黄酮生物合成的复杂反馈机制和精细分层调控ꎮ 调整ꎮ
苯丙烷途径的起始步骤往往会被下游代谢产物反 已有研究表明ꎬ红光可以调控药用植物次生
过来调节ꎬ同时也受相关酶的活性与含量变化影 代谢效应和机制ꎬ但不同物质的响应存在差异ꎮ
响ꎬ中间体( 如 t ̄CA) 可在供需匹配中促使上游转 一方面ꎬ红光能够激活光敏色素( Phy) 信号通路ꎬ
录回落ꎬ同时增强下游消耗ꎬ从而在转录层面呈现 通过 Phy 介导下游激素与转录因子网络调控黄酮
“上游 低 而 下 游 高” 的 动 态 平 衡 ( Wang et al.ꎬ 合成ꎮ 例如ꎬ蓝光和红光共同诱导的 R2R3 ̄MYB
2025)ꎮ 此外ꎬ苯丙烷代谢关键酶可在膜相关位点 转录因 子 FtMYB116 可 直 接 调 控 芦 丁 合 成 基 因