
Page 77 - 《广西植物》2020年第4期
P. 77
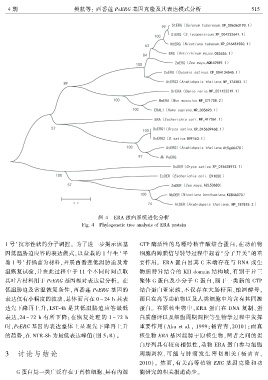
4 期 樊航等: 西番莲 PeERG 基因克隆及其表达模式分析 5 1 5
图 4 ERA 蛋白系统进化分析
Fig. 4 Phylogenetic tree analysis of ERA protein
1 号’抗寒性状的分子调控ꎮ 为了进一步揭示该基 GTP 酶活性的鸟嘌呤核苷酸结合蛋白ꎬ在动植物
因低温胁迫应答的表达模式ꎬ以盆栽的 1 年生‘ 平 细胞内跨膜信号转导过程中起着“分子开关” 的重
塘 1 号’扦插苗为材料ꎬ开展西番莲低温胁迫及常 要作用ꎮ ERA 蛋白因其 C 末端存在与 RNA 或生
温恢复试验ꎬ并在此过程中于 11 个不同时间点取 物膜特异结合的 KH domain 结构域ꎬ有别于异三
其叶片材料用于 PeERG 基因相对表达量分析ꎮ 在 聚体 G 蛋白及小分子 G 蛋白ꎬ属于一类新的 GTP
低温胁迫及常温恢复条件ꎬ西番莲 PeERG 基因的 结合蛋白亚家族ꎬ不仅存在大肠杆菌、酿酒酵母ꎬ
表达仅有小幅度的波动ꎬ总体而言在 0 ~ 24 h 其表 而且在高等动植物以及人类细胞中均含有其同源
达先下降再上升ꎬLST ̄4h 是其低温胁迫应答最低 蛋白ꎮ 在原核生物中ꎬERA 蛋白在 DNA 复制、蛋
表达ꎬ24 ~ 72 h 有所下降ꎻ在恢复处理的 1 ~ 72 h 白质翻译以及细胞周期调控等生物学过程中发挥
时ꎬPeERG 基因的表达整体上呈现先下降再上升 重要作用( Abu et al.ꎬ 1999ꎻ杨青青ꎬ2010)ꎻ而真
的趋势ꎬ在 NTR ̄8h 为最低表达峰值(图 5:B)ꎮ 核生物 ERA 基因起源于原核生物ꎬ两者之间的蛋
白序列具有较高相似性ꎬ动物 ERA 蛋白参与细胞
3 讨论与结论 周期调控、 可 能 与 肿 瘤 发 生 密 切 相 关 ( 杨 青 青ꎬ
2010)ꎮ 然而ꎬ有关高等植物 ERG 基因克隆和功
G 蛋白是一类广泛存在于真核细胞、具有内源 能研究的相关报道尚少ꎮ