
Page 80 - 《广西植物》2020年第8期
P. 80
8 期 姜楚等: 锥低磷胁迫响应基因的功能及代谢通路分析 1 1 3 5
传递链、脂质代谢、mRNA 加工、氧化还原过程、氧
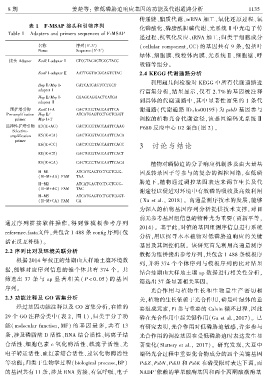
表 1 F ̄MSAP 接头和引物序列 化磷酸化、磷脂酰胆碱代谢、光系统Ⅱ中光电子传
Table 1 Adapters and primers sequences of F ̄MSAP
递过程、抗氧化反应、tRNA 加工ꎻ归类于细胞成分
名称 序列(5′ ̄3′) (cellular componentꎬCC) 的基因共有 9 条ꎬ包括叶
Name Sequence(5′ ̄3′)
绿体、细胞膜、线粒体内膜、光系统Ⅱ、细胞壁、呼
接头 Adapter EcoR I ̄adapter I CTCGTAGACTGCGTACC
吸链等组分ꎮ
EcoR I ̄adapterⅡ AATTGGTACGCAGTCTAC 2.4 KEGG 代谢通路分析
利用超几何检验对 KEGG 中所有代谢通路进
HapⅡ/ Msp I ̄ GATCATGAGTCCTGCT
行富集分析ꎬ结果显示ꎬ仅有 2.7% 的基因被注释
adapter I
HapⅡ/ Msp I ̄ CGAGCAGGACTCATGA 到具体的代谢通路中ꎬ其中显著性富集的 1 条代
adapterⅡ
预扩增引物 EcoR I+A GACTGCGTACCAATTCA 谢通路( 代谢通路 ID:ko00195) 为 psbD 基因参与
Pre ̄amplification HapⅡ/ ATCATGAGTCCTGCTCGGT
primer Msp I+T 调控的植物光合代谢途径ꎬ该基因编码光系统Ⅱ
选择性扩增引物 E3(E+AG) GACTGCGTACCAATTCAAG P680 反应中心 D2 蛋白(图 2)ꎮ
Selective ̄
amplification E5(E+CA) GACTGCGTACCAATTCACA
primer
3 讨论与结论
E6(E+CC) GACTGCGTACCAATTCACC
E8(E+CT) GACTGCGTACCAATTCACT
E9(E+GA) GACTGCGTACCAATTCAGA 植物对磷胁迫的分子响应机制涉及由大量基
H-M1 ATCATGAGTCCTGCTCGG ̄ 因及转录因子等参与的复杂的调控网络ꎬ在低磷
(H-M+AA) FAM TAA
胁迫下ꎬ植物通过调控基因表达来调节生长及代
H-M2 ATCATGAGTCCTGCTCGG ̄
(H-M+AC) FAM TAC
谢途径以促进对环境中有效磷的吸收及高效利用
H-M5 ATCATGAGTCCTGCTCGGT ̄
(H-M+CA) FAM CA (Xu et al.ꎬ 2018)ꎮ 高通量测序技术的发展ꎬ能够
为深入的植物基因序列分析提供技术支撑ꎬ对目
前无参考基因组信息的物种尤为重要( 贾新平等ꎬ
通过序 列 拼 接 软 件 操 作ꎬ 得 到 锥 模 板 参 考 序 列
2014)ꎮ 基于此ꎬ对锥的基因组测序信息进行系统
reference.fasta文件ꎬ共包含 1 488 条 contig 序列(包
分析ꎬ用以探寻木本植物对低磷胁迫响应的关键
括正反互补链)ꎮ
基因及其调控机制ꎮ 该研究首先利用高通量测序
2.2 序列比对及环境关联分析
数据为锥拼接出参考序列ꎬ共包含 1 488 条模板序
根据 2014 年校正的鼎湖山大样地土壤环境数
列ꎬ并将 374 个个体序列与模板序列的比对结果
据ꎬ能够对应序列信息的锥个体共有 374 个ꎮ 共
结合鼎湖山大样地土壤 ap 数据进行相关性分析ꎬ
筛选出 37 条 与 ap 显 著 相 关 ( P < 0. 05) 的 基 因
筛选出 37 条显著相关基因ꎮ
序列ꎮ
光合作用与植物生长和 生 物 量 生 产 密 切 相
2.3 功能注释及 GO 富集分析 关ꎬ植物的生长依赖于光合作用ꎬ磷是叶绿体的重
经过基因功能注释以及 GO 富集分析ꎬ在锥的 要组成元素ꎬPi 参与重要的 Calvin 循环过程ꎬ因此
29 个 GO 注释分类中( 表 2ꎬ 图 1)ꎬ归类于分子功 磷在光合作用中起关键作用(Gu et al.ꎬ2017)ꎮ 已
能( molecular functionꎬ MF) 的 基 因 最 多ꎬ 共 有 13 有研究表明ꎬ光合作用对低磷胁迫敏感ꎬ许多参与
条ꎬ涉及磷脂酶 D 活性、RNA 结合活性、钙离子结 光合作用的调控基因在受低磷胁迫时表达发生显
合活性、细胞色素 c 氧化酶活性、载流子活性、光 著变化(Slattery et al.ꎬ 2017)ꎮ 研究发现ꎬ大豆中
电子转运活性、血红素结合活性、过氧化物酶活性 编码光合过程中重要化合物成分的四个关键基因
等功能ꎻ归类于生物学过程(biological processꎬBP) PsbZ、PsbN、PsbH 和 PsbK 在磷受限时表达下调ꎬ而
的基因共有 11 条ꎬ涉及 RNA 剪接、有氧呼吸、电子 NADP 依赖的苹果酸酶基因和两个丙酮酸激酶基
+