
Page 114 - 《广西植物》2024年第12期
P. 114
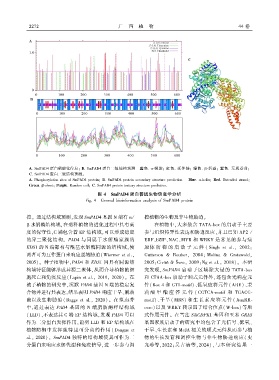
2 2 7 2 广 西 植 物 44 卷
A. SmPAD4 蛋白磷酸化位点ꎻ B. SmPAD4 蛋白二级结构预测 蓝色. α ̄螺旋ꎻ 红色. 延伸链ꎻ 绿色. β ̄折叠ꎻ 紫色. 无规卷曲ꎻ
C. SmPAD4 蛋白三级结构预测ꎮ
A. Phosphorylation sites of SmPAD4 proteinꎻ B. SmPAD4 protein secondary structure prediction Blue. α ̄helixꎻ Red. Extended strandꎻ
Green. β ̄sheetꎻ Purple. Random coilꎻ C. SmPAD4 protein tertiary structure prediction.
图 4 SmPAD4 蛋白普通生物信息学分析
Fig. 4 General bioinformatics analysis of SmPAD4 protein
控ꎮ 通过结构域预测ꎬ发现 SmPAD4 基因 N 端有 α / 控植物的生物及非生物胁迫ꎮ
β 水解酶结构域ꎬ在菊科植物的进化过程中具有高 在植物中ꎬ大多数含 TATA ̄box 的启动子主要
度的保守性ꎬC 端包含着 EP 结构域ꎬ可以形成稳定 参与组织特异性表达和胁迫反应ꎬ并且已知 AP2 /
的异二聚化结构ꎮ PAD4 与同属于水解酶家族的 ERF、bZIP、NAC、MYB 和 WRKY 是常见的参与病
EDS1 的 N 端都有与酰基水解酶同源的结构域ꎬ使 原体 防 御 的 启 动 子 元 件 ( Singh et al.ꎬ 2002ꎻ
两者可为互作蛋白来响应逆境胁迫(Wiermer et al.ꎬ Gutterson & Reuberꎬ 2004ꎻ Molina & Grotewoldꎬ
2005)ꎮ 种子植物中ꎬPAD4 和 EDS1 因具有同源结 2005ꎻCiváň & Švecꎬ 2009ꎻNg et al.ꎬ 2018)ꎮ 本研
构域特征能够形成异源二聚体ꎬ从而介导植物的细 究发现ꎬSmPAD4 启动子区域除大量的 TATA ̄box
胞死亡和免疫反应(Lapin et al.ꎬ 2019ꎬ 2020)ꎮ 在 和 CTAA ̄box 启动子顺式元件外ꎬ还包含光响应元
被子植物的研究中ꎬ沉默 PAD4 基因 N 端的稳定复 件(Box 4 和 GT1 ̄motif)、低氧应答元件( ARE)、茉
合物并进行共表达ꎬ结果表明 PAD4 响应干旱、脱落 莉 酸 甲 酯 应 答 元 件 ( CGTCA ̄motif 和 TGACG ̄
酸以及生物胁迫( Baggs et al.ꎬ 2020)ꎮ 在拟南芥 motif)、干旱 ( MBS) 和 生 长 素 应 答 元 件 ( AuxRR ̄
中ꎬ通过表达 PAD4 基因的 N 端脂肪酶样结构域 core)以及 WRKY 转录因子结合位点(W ̄box) 等顺
(LLD)ꎬ不表达其 C 端 EP 结构域ꎬ发现 PAD4 可以 式作用元件ꎮ 在雪莲 SikCDPK1 基因和玉米 GRAS
作为二分蛋白发挥作用ꎬ说明 LLD 和 EP 结构域在 基因家族启动子的研究中均包含了光信号、低氧、
植物防御中发挥独特且可分离的作用( Dongus et 干旱、生长素和 MeJA 相关的顺式元件从而参与植
al.ꎬ 2020)ꎮ SmPAD4 独特的结构域使其可作为二 物的生长发育和调控生物与非生物胁迫响应( 史
分蛋白来响应水解代谢和免疫信号ꎬ进一步参与调 光珍等ꎬ2022ꎻ吴占清等ꎬ2024)ꎬ与本研究结果一