
Page 90 - 《广西植物》2025年第3期
P. 90
4 7 0 广 西 植 物 45 卷
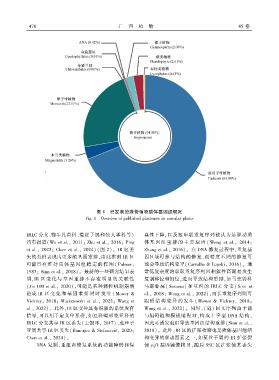
图 1 已发表的维管植物质体基因组概况
Fig. 1 Overview of published plastomes in vascular plants
IRLC 分支、牻牛儿苗科、菟丝子属和仙人掌科等) 真性下降ꎬ以及短串联重复序列被认为是驱动质
均有报道(Wu et al.ꎬ 2011ꎻ Zhu et al.ꎬ 2016ꎻ Ping 体基 因 组 重 排 的 主 要 原 因 ( Weng et al.ꎬ 2014ꎻ
et al.ꎬ 2022ꎻ Chen et al.ꎬ 2024) ( 图 2)ꎮ IR 区丢 Zhang et al.ꎬ 2016)ꎮ 在 DNA 修复过程中ꎬ重复基
失的类群表现出更多的基因重排ꎬ由此推测 IR 区 因区域可参与结构的修复ꎬ而精度不同的修复可
可能具有 维 持 质 体 基 因 组 稳 定 的 作 用 ( Palmerꎬ 能会导致结构变异(Carvalho & Lupskiꎬ 2016)ꎮ 通
1983ꎻ Sinn et al.ꎬ 2018)ꎮ 最新的一些研究结果表 常低复杂度的串联重复序列因相似性高而易发生
明ꎬIR 区 变 化 与 基 因 重 排 不 存 在 明 显 的 关 联 性 复制移位和倒位ꎬ进而导致结构重排ꎬ如马兜铃科
(Jin DM et al.ꎬ 2020)ꎬ可能是某种调控机制崩溃 马蹄香属( Saruma) 和豆科的 IRLC 分支( Sinn et
造成 IR 区 变 化 和 基 因 重 排 同 时 发 生 ( Mower & al.ꎬ 2018ꎻ Wang et al.ꎬ 2022)ꎬ而长重复序列则可
Vickreyꎬ 2018ꎻ Maciszewski et al.ꎬ 2022ꎻ Wang et 阻碍 结 构 变 异 的 发 生 ( Mower & Vickreyꎬ 2018ꎻ
al.ꎬ 2022)ꎮ 此外ꎬIR 区变异具有很强的系统发育 Wang et al.ꎬ 2022)ꎮ 同时ꎬ( 近) 回文序列由于能
信号ꎬ可以用于定义单系群ꎬ如豆科蝶形花亚科的 与编码链和模板链配对ꎬ构成十字型 DNA 结构ꎬ
IRLC 分支共享 IR 区丢失(王银环ꎬ 2017)、菟丝子 因此可诱发重组导致基因组结构重排( Sinn et al.ꎬ
亚属共享 IR 区丢失( Banerjee & Stefanovic'ꎬ 2020ꎻ 2018)ꎮ 此外ꎬIR 区的扩张收缩也是质体基因组结
Chen et al.ꎬ 2024)ꎮ 构变异的驱动因素之一ꎬ如菟丝子属的 IR 扩张促
DNA 复制、重组和修复系统的功能障碍和保 使 ycf1 基因镜像拷贝ꎬ随后 SSC 区扩张使其丢失