
Page 104 - 《广西植物》2025年第8期
P. 104
1 4 7 0 广 西 植 物 45 卷
有生理特性ꎬ由于基因组和遗传特性相对稳定ꎬ即
使环境因素发生变化ꎬ它们的高温适应性仍然表现
出一致性ꎮ 以往的研究也支持这一观点ꎬ认为植物
在面对环境变化时可能存在“气候记忆”ꎬ并通过实
验进一步验证了植物光合作用的温度适应能力受
到原产地气候条件的长期影响( Lin et al.ꎬ 2013ꎻ
Peltier et al.ꎬ 2022)ꎮ 综上认为ꎬ3 种栎类光合系统
温度适应性具有协调和一致性ꎬ这一结论有助于我
们更好地理解植物对高温环境的适应机制ꎮ
另外ꎬ本研究认为 3 种栎类光合系统高温适应
性与其 地 理 分 布 显 著 相 关ꎬ 这 一 发 现 与 周 帅 等
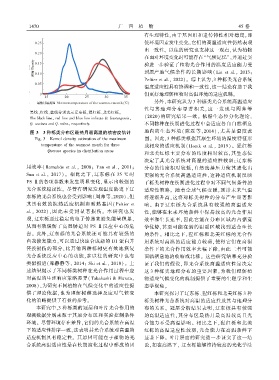
黑线、红线、蓝线分别表示辽东栎、猩红栎、北美红栎ꎮ
(2020)的研究结果一致ꎮ 依据生态位分化理论ꎬ
The black lineꎬ red line and blue line indicate Q. liaotungensisꎬ
不同物种在长期进化过程中会适应各自自然栖息
Q. coccinea and Q. rubraꎬ respectively.
图 3 3 种栎类分布区最热月最高温的核密度估计 地内的生态环境( 陈磊等ꎬ2014)ꎬ尤其是温度范
Fig. 3 Kernel density estimation of the maximum 围ꎮ 因此ꎬ3 种栎类根据其原生环境的温度特征形
temperature of the warmest month for three 成相应的适应机制( Horak et al.ꎬ 2019)ꎮ 猩红栎
Quercus species in distribution areas
和北美红栎主要分布的纬度相对较高ꎬ其生态位
决定了其光合系统对高温的适应性较弱ꎻ辽东栎
用效率( Barnabás et al.ꎬ 2008ꎻ Yan et al.ꎬ 2011ꎻ 分布的纬度相对较低ꎬ自然选择压力使其进化出
Sun et al.ꎬ 2017)ꎮ 相比之下ꎬ辽东栎在 35 ℃ 时 更强的光合系统高温适应性ꎬ这种适应机制反映
PS Ⅱ的各项参数未发生明显变化ꎬ显示出较强的 了栎类树种在长期进化过程中对不同气候条件的
光合系统稳定性ꎮ 尽管有研究发现温度胁迫下辽 适应性策略ꎮ 随着全球气候变暖ꎬ预计未来气温
东栎的光合系统也会受到影响( 周帅等ꎬ2020)ꎬ但 将逐渐升高ꎬ这将对栎类树种的分布产生显著影
其具有效的抗热适应机制和耐热基因( Peltier et 响ꎮ 由于辽东栎光合系统具有较强的高温适应
al.ꎬ 2022)ꎬ因 此 未 受 到 显 著 损 伤ꎮ 本 研 究 也 发 性ꎬ能够在未来环境条件中保持较高的光合作用
现ꎬ辽东栎通过稳定的电子传递来避免能量积累ꎬ 效率和生长速率ꎬ因此它能在分布区域内占据竞
从而有效缓解了高温胁迫对 PS Ⅱ反应中心的危 争优势ꎬ甚至可能在新的温暖区域找到适合生长
害ꎮ 此外ꎬ辽东栎的光合系统还可能具有较强的 的条件ꎮ 相比之下ꎬ猩红栎和北美红栎的光合作
自我修复能力ꎬ可以通过快速合成新的 D1 蛋白并 用系统对高温的适应能力较弱ꎬ使得它们在高温
替换损伤的部分ꎬ比其他两种栎树更有效地恢复 条件下的光合作用效率大幅下降ꎬ由此二者可能
光合系统反应中心的功能ꎬ在以往的研究中也有 面临栖息地的收缩或北移ꎮ 这些研究结果充分验
类似报道(郑静静等ꎬ 2014ꎻ Shi et al.ꎬ 2018)ꎮ 上 证了我们的假设ꎬ即光合系统高温适应性是决定
述结果揭示了不同栎类树种在光合作用过程中应 这 3 种栎类地理分布的重要因素ꎬ为我们理解植
对高温的生理响应策略差异( Takahashi & Murataꎬ 物适应气候变化的机制提供了重要的生理学和生
2008)ꎬ为研究不同植物在气候变化中的适应性提 态学视角ꎮ
供了理论依据ꎬ也为理解树种选择及应对气候变 本研究探讨了辽东栎、猩红栎和北美红栎 3 种
化的策略提供了有益的参考ꎮ 栎类树种光合系统对高温的适应性及其与地理分
本研究中ꎬ3 种栎类的冠层和叶片光合作用的 布的关系ꎮ 冠层分析结果表明ꎬ辽东栎具有较强
观测数据分别来源于其原分布区和实验控制条件 的高温适应性ꎬ其分布区最热月最高温较高且光
环境ꎮ 尽管环境存在差异ꎬ它们的光合系统在高温 合能力不受高温影响ꎮ 相比之下ꎬ猩红栎和北美
下的适应性排序一致ꎬ这表明其光合系统对高温的 红栎的高温适应性较弱ꎬ光合能力在高温条件下
适应机制具有稳定性ꎮ 其原因可能在于植物的光 显著下降ꎮ 叶片层面的研究进一步证实了这一结
合系统高温适应性是在长期演化过程中形成的固 论ꎬ如在高温下ꎬ 辽东栎能够维持较高的光化学活