
Page 137 - 《广西植物》2025年第8期
P. 137
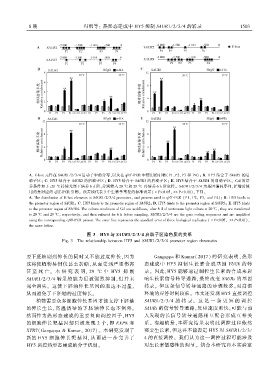
8 期 冯雨等: 热形态建成中 HY5 抑制 SAUR1 / 2 / 3 / 4 的转录 1 5 0 3
A. E ̄box 元件在 SAUR1 / 2 / 3 / 4 启动子中的分布ꎬ以及在 qRT ̄PCR 中使用的引物(P1、P2、P3 和 P4)ꎻ B. HY5 结合于 SAUR1 的启
动子区ꎻ C. HY5 结合于 SAUR2 的启动子区ꎻ D. HY5 结合于 SAUR3 的启动子区ꎻ E. HY5 结合于 SAUR4 的启动子区ꎮ Col 的培
养条件如下:20 ℃ 持续光照下培养 6 d 后ꎬ分别转入 20 ℃ 和 29 ℃ 再培养 6 h 后取样ꎮ SAUR1 / 2 / 3 / 4 为基因编码序列ꎬ扩增时使
用的是对应的 qRT ̄PCR 引物ꎮ 误差线代表 3 个生物学重复的标准误差(∗P<0.05ꎬ∗∗P<0.01)ꎬ下同ꎮ
A. The distribution of E ̄box elements in SAUR1/ 2/ 3/ 4 promotersꎬ and primers used in qRT ̄PCR (P1ꎬ P2ꎬ P3ꎬ and P4)ꎻ B. HY5 binds to
the promoter region of SAUR1ꎻ C. HY5 binds to the promoter region of SAUR2ꎻ D. HY5 binds to the promoter region of SAUR3ꎻ E. HY5 binds
to the promoter region of SAUR4. The culture conditions of Col are as follows: after 6 d of continuous light culture at 20 ℃ꎬ they are transferred
to 20 ℃ and 29 ℃ꎬ respectivelyꎬ and then cultured for 6 h before sampling. SAUR1/ 2/ 3/ 4 are the gene coding sequences and are amplified
using the corresponding qRT ̄PCR primer. The error line represents the standard error of three biological replicates (∗P<0.05ꎬ ∗∗P<0.01)ꎬ
the same below.
图 3 HY5 与 SAUR1/ 2/ 3/ 4 启动子区染色质的关系
Fig. 3 The relationship between HY5 and SAUR1/ 2/ 3/ 4 promoter region chromatin
要下胚轴剧烈伸长的同时又不能过度伸长ꎬ因为 Gangappa 和 Kumar( 2017) 的研究表明ꎬ热形
这将使植物易倒伏甚至折断ꎬ从而受到严重伤害 态建成中 HY5 抑制生长素合成基因 YUC8 的转
甚 至 死 亡ꎮ 本 研 究 表 明ꎬ 29 ℃ 中 HY5 抑 制 录ꎮ 因此ꎬHY5 能够通过调控生长素的合成来影
SAUR1 / 2 / 3 / 4 转录的能力虽被强烈抑制ꎬ但并未 响生长素信号转导通路ꎬ最终改变 SAURs 的基因
完全消失ꎮ 这使下胚轴伸长基因的表达不过量ꎬ 转录ꎮ 但这条信号转导通路的步骤较多ꎬ对高温
从而避免了下胚轴的过度伸长ꎮ 环境的应答时间较长ꎮ 本文还发现 HY5 直接调控
植物需要众多细胞伸长基因才能支撑下胚轴 SAUR1 / 2 / 3 / 4 的 转 录ꎮ 这 是 一 条 更 短 的 调 控
的伸长生长ꎬ高温诱导的下胚轴伸长也不例外ꎮ SAURs 的信号转导通路ꎬ反应速度更快ꎬ可能与前
然而作为热形态建成的重要负向调控因子ꎬHY5 人发现的长信号转导通路相互配合形成互补关
的细胞伸长靶基因却只被发现 2 个ꎬ即 EXP8 和 系ꎮ 有趣的是ꎬ本研究结果表明此调控过程依然
XTR7(Gangappa & Kumarꎬ 2017)ꎮ 本研究发现了 需要生长素ꎬ但这并不能否定 HY5 对 SAUR1 / 2 / 3 /
新的 HY5 细 胞 伸 长 靶 基 因ꎬ从 而 进 一 步 完 善 了 4 的直接调控ꎮ 我们认为这一调控过程可能涉及
HY5 调控热形态建成的分子机制ꎮ 对生长素敏感性的调节ꎮ 结合本研究和本实验室