
Page 66 - 《广西植物》2025年第8期
P. 66
1 4 3 2 广 西 植 物 45 卷
命以 应 对 增 温 引 起 的 环 境 变 化 ( Arroyo et al.ꎬ
2013)ꎮ 植物的雄性期与雌性期在发育过程中ꎬ对
温度的要求有所不同ꎮ 相关研究显示ꎬ雄性器官
发育的最适温度范围相对较窄ꎬ并且整体温度要
求偏低ꎬ而雌 性 器 官 的 发 育 则 与 之 相 反 ( Sage et
al.ꎬ 2015)ꎮ 正因如此ꎬ增温后蓝翠雀花雄性期的
缩短相较于雌性期更为显著ꎮ 与此同时ꎬ单花水
平的改变必然会对植物个体的开花持续时间产生
影响ꎬ因此ꎬ增温之后单花寿命的缩短是蓝翠雀花
单株开花持续时间缩短的一个关键原因ꎮ 在传粉
者稀少的高寒环境中ꎬ花寿命缩短一方面会减少
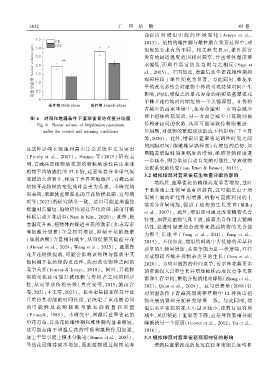
图 6 对照和增温条件下蓝翠雀花的花蜜分泌量 整个植株的花展示ꎬ另一方面会减少开花期间被
Fig. 6 Nectar volume of Delphinium caeruleum 传粉者访问的次数ꎬ从而可能导致传粉限制进一
under the control and warming conditions 步加剧ꎬ对植物的繁殖成功造成不利影响( 王玉贤
等ꎬ2020)ꎮ 此外ꎬ增温后蓝翠雀花两性时期之间
的间隔时间( 即雌雄异熟程度) 有缩短的趋势ꎬ如
且这种影响在极地和高山生态系统中尤为突出
果随着增温时间和幅度的增加ꎬ雌雄异熟程度进
(Prevéy et al.ꎬ 2017)ꎮ Sounan 等 ( 2019) 研 究 表
一步减小ꎬ则会增加自动自交的可能性ꎬ导致植物
明ꎬ青藏高原植物始花期的物候敏感性高达全球
交配系统的转变(van Etten & Brunetꎬ 2013)ꎮ
植物平均敏感性的 37.8 倍ꎬ这意味着在全球气候
3.2 模拟增温对蓝翠雀花生物量分配的影响
变暖的大背景下ꎬ相较于世界其他地区ꎬ青藏高原
增温后ꎬ蓝翠雀花的植株高度显著增加ꎬ茎叶
植物开花物候的变化或许会更为迅速ꎮ 本研究结
干重和地上生物量也有所提高ꎬ这可能是由于增
果表明ꎬ增温使蓝翠雀花的开花物候提前ꎬ这与魏
温使土壤的矿化作用增强ꎬ植物可直接利用的土
瑶等(2022)的研究结果一致ꎮ 原因可能是增温使
壤养分含量增加ꎬ促进了植物的生长发育( Miller
低温时长缩短ꎬ植物更快通过春化阶段ꎬ提前打破
et al.ꎬ 2007)ꎮ 此外ꎬ增温还可通过改变植物光合
休眠启动开花进程(Nam & Kimꎬ 2020)ꎮ 此外ꎬ随
特性ꎬ如促进植物气孔开放ꎬ提高光合作用关键酶
着温度升高ꎬ植物体内促进开花的激素( 如赤霉素
活性ꎬ促进叶绿素的合成等来提高植物的光合能
和细胞分裂素) 含量相对增加ꎬ抑制开花的激素
力和 生 长 速 率 ( Yang et al.ꎬ 2011ꎻ Fang et al.ꎬ
(如脱落酸) 含量相对减少ꎬ从而促使其提前开花
2015)ꎮ 不仅如此ꎬ增温后群落中占优势的禾草和
(Ahmad et al.ꎬ 2024ꎻ Wang et al.ꎬ 2025)ꎮ 蓝翠雀
莎草的生物量增加ꎬ其竞争能力进一步增强ꎬ往往
花开花物候提前ꎬ可能会影响该物种与群落中其
形成郁闭环境并抑制杂类草的生长( Chen et al.ꎬ
他同期开花植物的重叠性ꎬ从而改变物种之间的 2020)ꎮ 为应对激烈的种间竞争ꎬ蓝翠雀花将更多
竞争关系(Forrest & Inoiyeꎬ 2010)ꎮ 同时ꎬ开花物 的资源投入营养生长并增加植株高度以竞争光资
候的变化还可能打破植物与传粉者之间的同步
源和生存空间ꎬ繁殖分配就相应降低(Zhang et al.ꎬ
性ꎬ从而导致传粉失败( 肖宜安等ꎬ2015ꎻ施雨含 2021ꎻ Quan et al.ꎬ 2024)ꎮ 这与曹素珍(2018) 针
等ꎬ2021ꎻ王文等ꎬ2023)ꎮ 花寿命是指花保持开放 对增温条件下青藏高原高寒草甸中 12 种高山植
且维持其功能的时间长度ꎬ它决定了该花被访问 物开展的繁殖分配研究结果一致ꎮ 与此同时ꎬ增
的可 能 性 及 花 粉 接 收 与 散 布 的 数 量 和 质 量 温后蓝翠雀花的花大小显著减小ꎬ花数目也有所
(Primackꎬ 1985)ꎮ 本研究中ꎬ增温后蓝翠雀花的 减少ꎬ从而使花干重显著下降ꎬ这是导致繁殖分配
单花寿命ꎬ以及花的雄性期和雌性期均显著缩短ꎮ 降低的另一个原因(Hoover et al.ꎬ 2012ꎻ Yu et al.ꎬ
这可能是由于增温后花的呼吸和蒸腾作用加强ꎬ 2016)ꎮ
加上干旱引起土壤水分胁迫(Dunne et al.ꎬ 2003)ꎬ 3.3 模拟增温对蓝翠雀花花部特征的影响
导致花冠维持成本增加ꎬ因此植物通过缩短花寿 增温后蓝翠雀花的花冠直径和蜜距长度均显