
Page 117 - 《广西植物》2026年第4期
P. 117
4 期 高建有等: 叶片水分亏缺对猕猴桃细菌性溃疡病抗性的影响 6 7 5
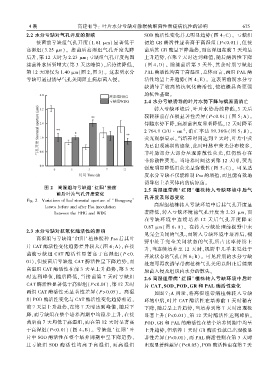
2.2 水分亏缺对气孔开度的影响 SOD 酶活性变化并无明显趋势( 图 4:C)ꎮ 亏缺组
接菌前亏缺组气孔开度(1.61 μm) 显著低于 初始 GR 酶活性显著高于高湿组( P<0.01)ꎬ但接
高湿组(3.25 μm)ꎮ 接菌后高湿组气孔开度先降 菌后其 GR 酶呈下降趋势ꎬ而高湿组在前 7 天均呈
后升ꎬ第 12 天时为 2.23 μmꎻ亏缺组气孔开度先因 上升趋势ꎬ在第 7 天时达到峰值ꎬ随后酶活性下降
接菌补水短暂增大(第 3 天达峰值)ꎬ后持续降低ꎬ (图 4:D)ꎮ 除接菌后第 3 天外ꎬ其余时期亏缺组
第 12 天时仅为 1.40 μm(图 2ꎬ图 3)ꎮ 这表明水分 PAL 酶活性均高于高湿组ꎬ总体而言ꎬ两组 PAL 酶
亏缺可通过诱导气孔关闭阻止病原菌入侵ꎮ 活性均呈上升趋势( 图 4:E)ꎮ 这表明前期水分亏
缺诱导了较高的抗氧化酶活性ꎬ使植株具备更强
的抗性基础ꎮ
2.4 水分亏缺诱导的叶片水势下降与病原菌消亡
转入亏缺环境后ꎬ叶片水势持续降低ꎬ3 天后
较转移前存在极显著性差异( P<0.01) ( 图 5:A)ꎮ
伴随水势下降ꎬ病原菌密度显著降低ꎬ12 天时降至
 ̄2
2 764.9 CFUcm ꎬ消亡率达 99.38% ( 图 5:B)ꎮ
荧光观察显示ꎬ当培养时间达到 7 天时ꎬ叶片中荧
光已出现减弱的迹象ꎬ此时叶脉中荧光分布较多ꎬ
非叶脉部分大部分呈现弥散性荧光ꎬ但仍然存在
非弥散性荧光ꎮ 当培养时间达到第 12 天时ꎬ荧光
强度明显降低且荧光呈弥散性( 图 5:C)ꎮ 可见适
度水分亏缺不仅能抑制 Psa 的增殖ꎬ而且能有效地
诱导宿主杀灭体内的病原菌ꎮ
图 2 高湿组与亏缺组‘红阳’接菌 2.5 高湿组带病‘ 红阳’ 植株转入亏缺环境中后气
前后叶片气孔开度变化
孔开度及形态变化
Fig. 2 Variations of leaf stomatal aperture of ‘Hongyang’
高湿组植株转入亏缺环境中后其气孔开度显
leaves before and after Psa inoculation
著降低ꎬ转入亏缺环境前气孔开度为 2.23 μmꎬ而
between the HHG and WDG
在亏缺 环 境 中 连 续 培 养 12 天 后 气 孔 开 度 降 至
0.87 μm( 图 6:A)ꎮ 在转入亏缺处理前视野中未
2.3 水分亏缺对抗氧化酶活性的影响
见完全关闭的气孔ꎬ而转入亏缺环境中培养后ꎬ视
高湿组与亏缺组‘ 红阳’ 植株接种 Psa 后其叶
野中处于完全关闭状态的气孔所占比率持续上
片 CAT 酶活性变化趋势差异较大( 图 4:A)ꎬ在接
升ꎬ当连续培养至 12 天时ꎬ视野中几乎未见处于
菌前亏缺组 CAT 酶活性显著高于高湿组( P < 0.
开放状态的气孔(图 6:B)ꎮ 可见后期的水分亏缺
01)ꎬ但接菌后亏缺组 CAT 酶活性呈下降趋势ꎬ而
处理可再次诱导带病植株气孔关闭以阻止后续病
高湿组 CAT 酶活性在前 3 天呈上升趋势ꎬ第 3 天
原菌入侵及组织内水分的散失ꎮ
时达到峰值ꎬ随后降低ꎬ当接菌第 7 天时亏缺组 2.6 高湿组带病‘ 红阳’ 植株转入亏缺环境中后叶
CAT 酶活性显著低于高湿组(P<0.01)ꎬ第 12 天时 片 CAT、SOD、POD、GR 和 PAL 酶活性变化
两组 CAT 酶活性无显著性差异( P>0.05)ꎮ 高湿 如图 7:A 所示ꎬ将高湿组带病植株转入亏缺
组 POD 酶活性变化与 CAT 酶活性变化趋势相近ꎬ 环境中后ꎬ叶片 CAT 酶活性在培养前 1 天时稍有
前 7 天呈上升趋势ꎬ在第 7 天时达到峰值ꎬ随后下 下降ꎬ随后呈上升趋势ꎬ当培养到第 7 天时出现极
降ꎬ而亏缺组在整个培养周期中均稳步上升ꎬ在接 显著上升(P<0.01)ꎬ第 12 天时酶活性达到峰值ꎮ
菌后前 7 天均低于高湿组ꎬ而在第 12 天时显著高 POD、GR 和 PAL 的酶活性在整个培养周期中均呈
于高湿组(P<0.01) ( 图 4:B)ꎮ 亏缺组‘ 红阳’ 叶 上升趋势ꎬ但培养 1 天时 GR 酶活性就已出现极显
片中 SOD 酶活性在整个培养周期中呈下降趋势ꎬ 著性差异(P<0.01)ꎬ而 PAL 酶活性则在第 3 天时
且亏缺 组 SOD 酶 活 性 均 高 于 高 湿 组ꎬ 而 高 湿 组 出现显著性差异(P<0.05)ꎬPOD 酶活性也在第 7 天