
Page 120 - 《广西植物》2026年第4期
P. 120
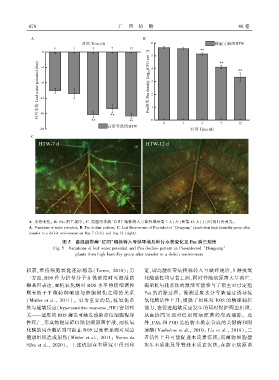
6 7 8 广 西 植 物 46 卷
A. 水势变化ꎻ B. Psa 消亡规律ꎻ C. 高湿组带病‘红阳’植株转入亏缺环境后第 7 天(左)和第 12 天(右)时的叶片荧光ꎮ
A. Variations of water potentialꎻ B. Psa decline patternꎻ C. Leaf fluorescence of Psa ̄infected ‘Hongyang’ plants from high humidity group after
transfer to a deficit environment on Day 7 (left) and Day 12 (right).
图 5 高湿组带病‘红阳’植株转入亏缺环境后叶片水势变化及 Psa 消亡规律
Fig. 5 Variations of leaf water potential and Psa decline pattern in Psa ̄infected ‘Hongyang’
plants from high humidity group after transfer to a deficit environment
积累ꎬ维持细胞氧化还原稳态( Torresꎬ 2010)ꎻ另 证ꎬ即高湿组带病植株转入亏缺环境后ꎬ5 种抗氧
一方面ꎬROS 作为信号分子在低浓度时可激活防 化酶活性均显著上调ꎬ同时伴随病原菌大量消亡ꎬ
御基因表达ꎬ而抗氧化酶对 ROS 水平的精细调控 提示抗氧化系统的激活可能参与了宿主对已定殖
则有助于平衡防御响应与细胞损伤之间的关系 Psa 的清除过程ꎮ 推测适度水分亏缺通过诱导抗
(Mittler et al.ꎬ 2011)ꎮ 更为重要的是ꎬ抗氧化系 氧化酶活性上升ꎬ增强了植株对 ROS 的精细调控
统与超敏反应(hypersensitive responseꎬHR) 密切相 能力ꎬ在促进超敏反应发生的同时保护周边组织ꎬ
关———适度的 ROS 爆发可触发感染部位细胞程序 从而协同实现对已定殖病原菌的有效清除ꎮ 此
性死亡ꎬ形成物理屏障以限制病原菌扩散ꎬ而抗氧 外ꎬPAL 和 POD 是植物木质素合成的关键酶和限
化酶的同步激活则可防止 ROS 过度积累而对周边 速酶(Vanholme et al.ꎬ 2010ꎻ Liu et al.ꎬ 2018)ꎬ二
健康组织造成损伤( Mittler et al.ꎬ 2011ꎻ Nunes da 者活性上升可能促进木质素沉积ꎻ而植物细胞壁
Silva et al.ꎬ 2020)ꎮ 上述机制在本研究中得到印 次生木质化及导管处木质素沉积ꎬ在防止病原菌