
Page 164 - 《广西植物》2026年第4期
P. 164
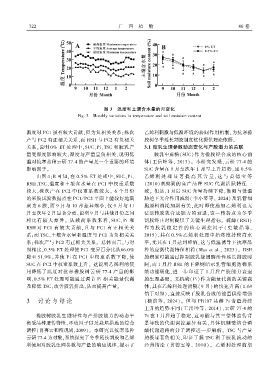
7 2 2 广 西 植 物 46 卷
图 3 温度和土壤含水量的月变化
Fig. 3 Monthly variations in temperature and soil moisture content
温度对 PC1 虽有较大贡献ꎬ但为负相关关系ꎻ株次 乙烯利刺激与低温环境的协同作用机制ꎬ为抗寒橡
产与 PC2 有正相关关系ꎬ而 RSH 与 PC2 有负相关 胶树冬季延长割胶制度优化提供理论依据ꎮ
关系ꎬ说明 0% ET 处理中ꎬSUC、Pi、TSC 和胶乳产 3.1 胶乳生理参数动态变化与产胶潜力的关联
量受温度影响较大ꎬ温度与产量呈负相关ꎬ说明低 胶乳中蔗糖(SUC) 作为橡胶烃合成的核心前
温对抗寒品种云研 77 ̄4 的产量是一个重要的环境 体(王岳坤等ꎬ 2013)ꎮ 本研究发现ꎬ云研 77 ̄4 的
影响因子ꎮ SUC 含量在 9 月至次年 1 月呈上升趋势ꎬ且 0.5%
由图 4:B 可知ꎬ在 0.5% ET 处理中ꎬSUC、Pi、 乙烯 利 处 理 显 著 提 高 其 含 量ꎬ 这 与 黄 德 宝 等
RSH、TSC、温度和土壤含水量在 PC1 中权重系数 (2010)观察到的高产品种 SUC 代谢活跃特征一
较大ꎬ株次产在 PC2 中权重系数较大ꎮ 6 个月份 致ꎮ 但是ꎬ1 月后 SUC 含量持续下降ꎬ推测与低温
的采胶试验数据点在 PC1 / PC2 平面上能较好地集 胁迫下光合作用减弱( 李小琴等ꎬ 2024) 及乳管细
聚为 6 簇ꎬ而 9 月与 10 月差异缩小ꎬ仅 9 月与 11 胞原料消耗加剧有关ꎬ此时即使施加乙烯利也无
月至次年 2 月显著分离ꎬ说明 9 月与其他月份之间 法逆转胶乳合成能力的衰退ꎬ这一转折点为冬季
相比 有 较 大 差 异ꎮ 从 载 荷 系 数 来 看ꎬ SUC、 Pi 和 割胶终止时间提供了关键生理指标ꎮ 硫醇( RSH)
RSH 对 PC1 有 较 大 贡 献ꎬ且 与 PC1 有 正 相 关 关 作为 胶 乳 稳 定 性 的 核 心 调 控 因 子 ( 史 敏 晶 等ꎬ
系ꎻ而 TSC、土壤含水量和温度与 PC1 为负相关关 2015)ꎬ其在 0.5%乙烯利处理中始终维持较高水
系ꎻ株次产与 PC2 有正相关关系ꎮ 总体而言ꎬ与对 平ꎬ尤其在 1 月达到峰值ꎬ这与低温诱导下抗寒品
照相比ꎬ0.5% ET 处理使 PC1 变异百分比从66.0% 种的应激代谢特征相符( Mao et al.ꎬ 2023)ꎮ RSH
降至 51.9%ꎬ并使 Pi 在 PC1 中权重系数下降ꎬ使 的积累可能通过抑制胶乳凝固酶活性延长排胶时
SUC 在 PC2 中权重系数上升ꎮ 这说明乙烯利的使 间ꎬ而 1 月后 RSH 的下降则暗示乳管细胞防御系
用降低了温度对抗寒橡胶树云研 77 ̄4 产量的影 统功能弱化ꎬ进一步印证了 1 月后产胶能力衰退
响ꎬ0.5% ET 处理可能通过调节 Pi 相关能量代谢 的生理基础ꎮ 无机磷( Pi) 作为能量代谢的关键载
及降低 TSCꎬ改善胶乳排出ꎬ从而提高产量ꎮ 体ꎬ其在乙烯利处理初期(9 月) 的快速升高(1.69
倍于对照)ꎬ直接反映了胶乳合成的能量供给增强
3 讨论与结论 (杨洪等ꎬ 2024)ꎮ 但与 PR107 品种 Pi 含量持续
上升的趋势不同( 王岳坤等ꎬ 2014)ꎬ云研 77 ̄4 的
橡胶树胶乳生理特性与产排胶能力的动态平 Pi 在 1 月后趋于稳定ꎬ这可能与其三倍体遗传背
衡受品种遗传特性、环境因子以及栽培措施的综合 景导致的代谢调控差异有关ꎬ具体机制需结合磷
调控(肖再云和校现周ꎬ2009)ꎮ 本研究以抗寒品种 酸代谢通路的分子调控进一步解析ꎮ TSC 与产量
云研 77 ̄4 为对象ꎬ系统探究了冬季延长割胶和乙烯 的极显著负相关ꎬ印证了低 TSC 利于胶乳流动的
利使用对胶乳生理参数与产量的响应规律ꎬ揭示了 经典结论 ( 黄德宝等ꎬ 2010) ꎮ 乙烯利处理前期