
Page 52 - 《广西植物》2026年第4期
P. 52
6 1 0 广 西 植 物 46 卷
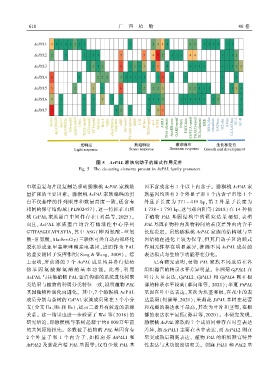
图 5 AcPAL 家族启动子的顺式作用元件
Fig. 5 The cis ̄acting elements present in AcPAL family promoters
串联重复与片段复制是驱动猕猴桃 AcPAL 家族数 因不含或含有 1 个以上内含子ꎮ 猕猴桃 AcPAL 家
量扩张的主要因素ꎮ 猕猴桃 AcPAL 家族编码的蛋 族基因均具有 2 个外显子和 1 个内含子且第 1 个
白不仅基序的排列顺序和数量高度一致ꎬ还含有 外显子长度 为 371 ~ 419 bpꎬ第 2 外 显 子 长 度 为
相同的保守结构域( PLN02457)ꎬ这一特征在山核 1 738 ~ 1 750 bpꎬ这与郝向阳等(2018) 在 14 种被
桃 CcPAL 家族蛋白中同样存在( 刘晨等ꎬ2025)ꎮ 子植物 PAL 基 因 结 构 中 的 研 究 结 果 相 似ꎬ 表 明
而且ꎬAcPAL 家 族 蛋 白 均 含 有 酶 活 性 中 心 序 列 PAL 基因在物种内及物种间的长度差异由内含子
GTITASGDLVPLSYIAꎬ其中 ASG( 即丙氨酸 - 丝氨 长度决定ꎮ 虽然猕猴桃 AcPAL 家族的结构域与基
酸-甘氨酸ꎬAla ̄Ser ̄Gly)三联体可经自动内部环化 因结构在进化上较为保守ꎬ但其启动子区的顺式
脱水形成亚甲基咪唑酮亲电基团ꎬ进而作为 PAL 作用元件存在明显差异ꎬ推测不同 AcPAL 成员的
的重要辅因子发挥作用(Song & Wangꎬ 2009)ꎮ 综 表达模式与生物学功能存在分化ꎮ
上表明ꎬ所获得的 7 个 AcPAL 成员均具备行使植 已有研究表明ꎬ植物 PAL 家族不同成员在各
物 苯 丙 氨 酸 解 氨 酶 的 基 本 功 能ꎮ 此 外ꎬ 利 用 组织器官的转录水平差异明显ꎮ 非洲菊 GjPAL1 在
AcPAL 与其他植物 PAL 蛋白构建的系统进化树聚 叶片大量表达ꎬGjPAL2、GjPAL3 和 GjPAL4 则在根
类结果与植物的种属分类特征一致ꎬ说明植物 PAL 部的转录水平较高(郝向阳等ꎬ 2021)ꎻ半夏 PtPAL
基因随物种演化而进化ꎮ 其中ꎬ7 个猕猴桃 AcPAL 基因在叶中高表达ꎬ其次为块茎和根ꎬ在花中的表
成员分别与茶树的 CsPAL 家族成员聚在 3 个小分 达最弱(何潇等ꎬ2021)ꎻ茉莉花 JsPAL 基因在花蕾
支(分支 IIa、IIb 和 IIc)ꎬ提示二者具有较近的亲缘 和花瓣的表达水平最高ꎬ其次为叶片和茎部ꎬ在根
关系ꎮ 这一结果也进一步验证了 Wei 等(2018) 的 部的表达水平最低(孙君等ꎬ2020)ꎮ 本研究发现ꎬ
研究结论ꎬ即猕猴桃与茶树起源于约8 000万年前 猕猴桃 AcPAL 家族的 7 个成员同样存在时空表达
的共同原始祖先ꎮ 多数被子植物的 PAL 基因含有 差异ꎬ如 AcPAL1 主要在茎中表达ꎬ而 AcPAL2 则在
2 个外 显 子 和 1 个 内 含 子ꎬ 如 拟 南 芥 AtPAL1 和 果实成熟后期高表达ꎮ 植物 PAL 的组织器官特异
AtPAL2 及紫花苜蓿 PAL 基因等ꎻ仅有少数 PAL 基 性表达与其功能密切相关ꎮ 剑麻 PAL1 和 PAL2 基