
Page 47 - 《广西植物》2026年第4期
P. 47
4 期 冯新等: 猕猴桃 AcPAL 家族基因的鉴定及在果实成熟过程的表达 6 0 5
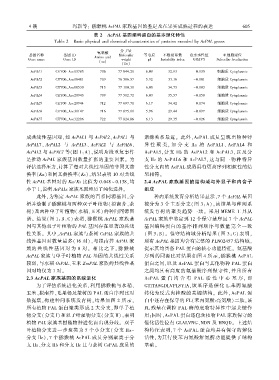
表 2 AcPAL 基因编码蛋白的基本理化特性
Table 2 Basic physical and chemical characteristics of proteins encoded by AcPAL genes
分子量
氨基酸
基因名称 基因 ID Molecular 等电点 不稳定系数 总亲水性值 亚细胞定位
Amino acid
Gene name Gene ID weight pI Instability index GRAVY Subcellar localization
(aa)
(Da)
AcPAL1 CEY00_Acc03785 706 77 044.20 6.09 32.93 -0.135 细胞质 Cytoplasmic
AcPAL2 CEY00_Acc08481 709 76 586.57 5.92 33.16 -0.101 细胞质 Cytoplasmic
AcPAL3 CEY00_Acc08530 713 77 100.31 6.00 34.73 -0.103 细胞质 Cytoplasmic
AcPAL4 CEY00_Acc20943 709 77 502.72 6.03 35.57 -0.150 细胞质 Cytoplasmic
AcPAL5 CEY00_Acc20944 712 77 697.71 6.17 34.42 -0.174 细胞质 Cytoplasmic
AcPAL6 CEY00_Acc30147 716 77 075.00 5.96 29.44 -0.097 细胞质 Cytoplasmic
AcPAL7 CEY00_Acc32206 722 77 824.86 6.13 29.35 -0.126 细胞质 Cytoplasmic
成共线性基因对ꎬ如 AcPAL1 与 AcPAL2、AcPAL1 与 亲缘关系最近ꎮ 此外ꎬAcPAL 成员呈现出物种特
AcPAL7、 AcPAL2 与 AcPAL3、 AcPAL2 与 AcPAL6、 异 性 聚 类ꎬ 如 分 支 IIa 的 AcPAL1、 AcPAL4 和
AcPAL2 与 AcPAL7 等(图 1:A)ꎬ说明片段重复事件 AcPAL5ꎬ分支 IIb 的 AcPAL2 和 AcPAL3ꎬ以及分
是推动 AcPAL 家族基因数量扩张的重要因素ꎮ 为 支 IIc 的 AcPAL6 和 AcPAL7ꎬ这与同一物种特异
评估选择压力ꎬ计算了每对共线性基因的非同义替 性分支内的 AcPAL 成员具有更高序列相似性的结
换率(Ka)和同义替换率(Ks)ꎬ结果表明 10 对共线 果相符ꎮ
性 AcPAL 基因对的 Ka / Ks 比值为 0.048 ~ 0.138ꎬ均 2.4 AcPAL 家族基因的结构域与外显子和内含子
小于 1ꎬ说明 AcPALs 家族基因经历了纯化选择ꎮ 组成
此外ꎬ为鉴定 AcPAL 家族的直系同源基因ꎬ分 种内系统发育分析结果显示ꎬ7 个 AcPAL 基因
析并绘制了猕猴桃与两种双子叶植物( 拟南芥、茶 被分为 3 个主要分支( 图 3:A)ꎬ该结果与种间系
树)及两种单子叶植物(水稻、玉米)的种间同源图 统发育树的聚类趋势一致ꎮ 采用 MEME 工具从
谱ꎮ 结果(图 1:B、C) 表明ꎬ猕猴桃 AcPAL 家族基 AcPAL 家族中鉴定到 12 个保守基序且 7 个 AcPAL
因与其他双子叶植物的 PAL 基因存在显著的共线 基因编码蛋白的基序排列顺序与数量完全一致
性关系ꎮ 其中ꎬAcPAL 家族与茶树 CsPAL 家族的共 (图 3:B)ꎮ 保守结构域分析结果( 图 3:C) 表明ꎬ
线性基因对数量最多(16 对)ꎬ与拟南芥 AtPAL 家 所有 AcPAL 基因均含有完整的 PLN02457 结构域ꎬ
族的 共 线 性 基 因 对 为 8 对ꎮ 相 比 之 下ꎬ 猕 猴 桃 提示其均具备 PAL 蛋白的核心功能特征ꎮ 氨基酸
AcPAL 家族与单子叶植物 PAL 基因的共线性关系 序列的同源比对结果如图 4 所示ꎬ猕猴桃 AcPAL
较弱ꎬ与水稻 OsPAL、玉米 ZmPAL 家族的共线性基 蛋白之间ꎬ以及 AcPAL 蛋白与其他物种 PAL 蛋白
因对均仅为 1 对ꎮ 之间均具有高度的氨基酸序列保守性ꎬ并且所有
2.3 AcPAL 家族基因的系统进化 AcPAL 蛋 白 均 含 有 PAL 活 性 中 心 基 序ꎬ 即
为了评估系统进化关系ꎬ利用猕猴桃与水稻、 GTITASGDLVPLSYIAꎬ该基序是催化 L ̄苯 丙 氨 酸
玉米、拟南芥、毛果杨及茶树的 PAL 蛋白序列比对 转化为反式肉桂酸的关键结构ꎮ 此外ꎬAcPAL 蛋
数据集ꎬ构建种间系统发育树ꎬ结果如图 2 所示ꎬ 白中还存在保守的 FL(苯丙氨酸 ̄亮氨酸)二肽ꎬ该
所有植物 PAL 蛋白聚类形成 2 大分支ꎬ即单子植 FL 残基在调控 PAL 酶的底物特异性中起关键作
物分支(分支 I)和双子叶植物分支(分支 II)ꎬ表明 用ꎻ同时ꎬAcPAL 蛋白还包含植物 PAL 家族保守的
植物 PAL 家族基因随物种进化而出现分歧ꎮ 双子 催化活性位点 GLALVNG、NDN 及 HNQDꎮ 上述结
叶植物分支进一步聚类为 5 个小分支( 分支 IIa- 构特征表明ꎬ7 个 AcPAL 蛋白均具有保守的催化
分支 IIe)ꎬ7 个猕猴桃 AcPAL 成员分别聚类于分 活性ꎬ为其行使苯丙氨酸解氨酶功能提供了结构
支 IIa、分支 IIb 和分支 IIc 且与茶树 CsPAL 成员的 基础ꎮ