
Page 50 - 《广西植物》2026年第4期
P. 50
6 0 8 广 西 植 物 46 卷
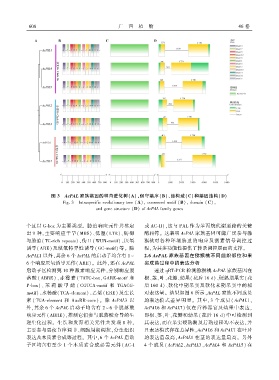
图 3 AcPAL 家族基因的种内进化树(A)、保守基序(B)、结构域(C)和基因结构(D)
Fig. 3 Intraspecific evolutionary tree (A)ꎬ conserved motif (B)ꎬ domain (C)ꎬ
and gene structure (D) of AcPAL family genes
个且以 G ̄box 为主要类型ꎮ 胁迫响应元件共鉴定 或 AC ̄II)ꎬ这与 PAL 作为苯丙烷代谢通路的关键
出 9 种ꎬ主要响应干旱( MBS)、低温( LTR)、防御 酶相符ꎮ 这表明 AcPAL 家族基因可能广泛参与猕
与胁迫( TC ̄rich repeats)、伤口( WUN ̄motif)、厌氧 猴桃对各种环境胁迫的响应及激素信号调控过
诱导( ARE) 及缺氧特异性诱导( GC ̄motif) 等ꎮ 除 程ꎬ为其多功能性提供了转录调控层面的支撑ꎮ
AcPAL1 以外ꎬ其余 6 个 AcPAL 的启动子均含有 1 ~ 2.6 AcPAL 家族基因在猕猴桃不同组织部位和果
6 个响应厌氧诱导元件(ARE)ꎮ 此外ꎬ还在 AcPAL 实成熟过程中的表达分析
启动子区检测到 10 种激素响应元件ꎬ分别响应脱 通过 qRT ̄PCR 检测猕猴桃 AcPAL 家族基因在
落酸 ( ABRE)、赤 霉 素 ( TATC ̄box、GARE ̄motif 和 根、茎、叶、花瓣、幼果(花后 16 d)、硬熟期果实(花
P ̄box)、 茉 莉 酸 甲 酯 ( CGTCA ̄motif 和 TGACG ̄ 后 160 d)、软化中期果实及软化末期果实中的相
motif)、水杨酸(TCA ̄element)、乙烯( ERE) 及生长 对表达量ꎮ 结果如图 6 所示ꎬAcPAL 家族不同成员
素 ( TGA ̄element 和 AuxRR ̄core)ꎮ 除 AcPAL3 以 的表达模式差异明显ꎮ 其中ꎬ3 个成员( AcPAL1、
外ꎬ其余 6 个 AcPAL 启动子均含有 2 ~ 6 个脱落酸 AcPAL6 和 AcPAL7) 仅在营养器官及幼果中表达ꎬ
响应元件(ABRE)ꎬ推测它们参与脱落酸介导的生 即根、茎、叶、花瓣和幼果( 花后 16 d) 中可检测到
理生化过程ꎮ 生长和发育相关元件共发现 8 种ꎬ 其表达ꎬ而在果实硬熟期及后熟过程均不表达ꎬ并
主要参与昼夜节律调节、细胞周期调控、分生组织 且表达模式存在差异性ꎬAcPAL6 和 AcPAL7 在叶片
表达及木质素合成等过程ꎮ 其中ꎬ6 个 AcPAL 启动 的表达量最高ꎬAcPAL1 在茎的表达量最高ꎮ 另外
子区均含有至少 1 个木质素合成必需元件( AC ̄I 4 个成员( AcPAL2、AcPAL3、AcPAL4 和 AcPAL5) 在