
Page 81 - 《广西植物》2026年第4期
P. 81
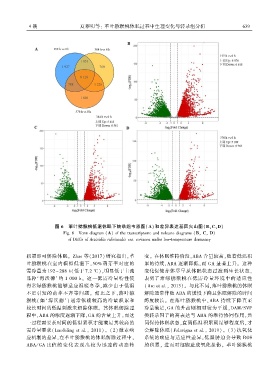
4 期 夏黎明等: 革叶猕猴桃休眠过程中生理变化与转录组分析 6 3 9
图 6 革叶猕猴桃低温休眠下转录组韦恩图(A)和差异表达基因火山图(B、C、D)
Fig. 6 Venn diagram (A) of the transcriptome and volcano diagrams (Bꎬ Cꎬ D)
of DEGs of Actinidia rubricaulis var. coriacea under low ̄temperature dormancy
积累即可解除休眠ꎮ Zhao 等(2017) 研究指出ꎬ革 变ꎮ 在休眠维持阶段ꎬABA 含量较高ꎬ随着低温积
叶猕猴桃在室内模拟低温下ꎬ50% 萌芽率对应的 累的持续ꎬABA 逐渐降低ꎬ而 GA 显著上升ꎮ 这种
需冷量为 192 ~ 288 h(低于 7.2 ℃ )ꎬ明显低于主流 变化促使芽体尽早从休眠状态过渡到生长状态ꎬ
品种‘海沃德’ 约 1 000 hꎮ 这一低需冷量特性使 表明了常绿猕猴桃在低需冷量环境中的适应性
得常绿猕猴桃能够适应温暖冬季ꎬ减少由于低温 (Arc et al.ꎬ 2013)ꎮ 与此不同ꎬ落叶猕猴桃的休眠
不足引发的萌芽不齐等问题ꎮ 相比之下ꎬ落叶猕 解除通常伴随 ABA 的缓慢下降且休眠解除的时间
猴桃( 如‘ 海沃德’) 通常依赖较高的冷量积累和 跨度较长ꎮ 在落叶猕猴桃中ꎬABA 持续下降直至
较长时间的低温刺激来解除休眠ꎮ 其休眠解除过 冷量满足ꎬGA 的升高则相对较为平缓ꎬDAM / SVP
程中ꎬABA 的浓度逐渐下降ꎬGA 的含量上升ꎬ而这 类转录因子的高表达与 ABA 的维持协同作用ꎬ共
一过程需要长时间的低温累积才能满足其较高的 同保持休眠状态ꎬ直到低温积累到足够程度后ꎬ才
需冷量要求(Luedeling et al.ꎬ 2011)ꎮ (2) 激素响 会解除休眠( Falavigna et al.ꎬ 2019)ꎮ (3) 抗氧化
应机制的差异ꎬ在革叶猕猴桃的休眠解除过程中ꎬ 系统的响应与适应性差异ꎬ低温胁迫会导致 ROS
ABA / GA 比值的 变 化 表 现 出 较 为 迅 速 的 动 态 转 的积累ꎬ 进而对细胞造成氧化损伤ꎮ 革叶猕猴桃