
Page 80 - 《广西植物》2026年第4期
P. 80
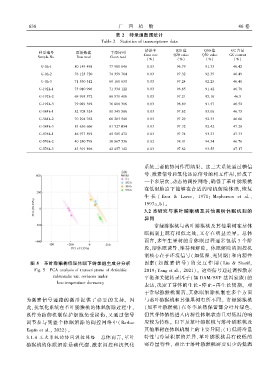
6 3 8 广 西 植 物 46 卷
表 2 转录组数据统计
Table 2 Statistics of transcriptome data
错误率 Q20 值 Q30 值 GC 含量
样品编号 原始数据 干净序列
Error rate Q20 value Q30 value GC content
Sample No. Raw read Clean read
(%) (%) (%) (%)
G ̄0h ̄1 80 149 498 77 980 846 0.03 96.79 91.33 46.42
G ̄0h ̄2 76 123 720 74 359 704 0.03 97.32 92.35 46.49
G ̄0h ̄3 71 530 312 69 160 630 0.03 97.24 92.23 46.48
G ̄192h ̄1 73 010 996 71 334 122 0.03 96.85 91.42 46.78
G ̄192h ̄2 69 995 572 68 571 406 0.03 97.21 92.16 46.5
G ̄192h ̄3 79 029 392 76 680 306 0.03 96.89 91.57 46.52
G ̄384h ̄1 82 708 324 80 345 586 0.03 97.62 93.06 46.75
G ̄384h ̄2 70 294 762 68 203 548 0.03 97.29 92.33 46.66
G ̄384h ̄3 83 656 666 81 527 054 0.03 97.32 92.42 47.28
G ̄576h ̄1 46 977 592 45 585 472 0.03 97.74 93.33 47.33
G ̄576h ̄2 40 240 798 38 867 536 0.02 98.01 94.34 46.76
G ̄576h ̄3 43 391 896 42 457 142 0.03 97.82 93.55 47.17
系统三者的协同作用结果ꎮ 这三大系统通过糖信
号、激素信号和氧化还原信号的相互作用ꎬ形成了
一个多层次、动态的调控网络ꎬ确保了革叶猕猴桃
在低温胁迫下能够在合适的时机解除休眠ꎬ恢复
生 长 ( Erez & Laveeꎬ 1971ꎻ Mcpherson et al.ꎬ
1997aꎬb)ꎮ
3.2 本研究与落叶猕猴桃及其他果树休眠机制的
异同
常绿猕猴桃与落叶猕猴桃及其他果树在芽休
眠机制上既有相似之处ꎬ又存在明显差异ꎮ 总体
而言ꎬ多年生果树的芽休眠过程通常包括 3 个阶
段ꎬ即休眠诱导、维持和解除ꎮ 休眠解除的调控机
制核心在于环境信号( 如低温、光周期) 和内源性
图 5 革叶猕猴桃低温休眠下转录组主成分分析 因素( 如 激 素 信 号) 的 交 互 作 用 ( Liu & Sherifꎬ
Fig. 5 PCA analysis of transcriptome of Actinidia 2019ꎻYang et al.ꎬ 2021)ꎮ 这些信号通过调控激素
rubricaulis var. coriacea under 平衡和关键转录因子( 如 DAM / SVP 基因家族) 的
low ̄temperature dormancy
表达ꎬ决定了芽体的生长-停止-再生长周期ꎮ 对
于常绿猕猴桃而言ꎬ其休眠解除机制在多个方面
为激素信号通路的激活提供了必要的支持ꎮ 因 与落叶猕猴桃和其他果树有所不同ꎮ 常绿猕猴桃
此ꎬ抗氧化系统在革叶猕猴桃的休眠解除过程中ꎬ (如革叶猕猴桃) 在冬季虽然保留部分叶片绿色ꎬ
既作为防御机制保护细胞免受损伤ꎬ又通过信号 但其芽体依然进入内源性休眠状态且对低温的响
调节参与到整个休眠解除的调控网络中( Barba ̄ 应较为特殊ꎮ 以下是革叶猕猴桃与落叶猕猴桃及
Espín et al.ꎬ 2022)ꎮ 其他果树在休眠机制上的主要异同:(1) 低需冷量
3.1.4 三大系统的协同调控网络 总体而言ꎬ革叶 特性与冷量积累的差异ꎬ革叶猕猴桃具有较低的
猕猴桃的休眠解除是碳代谢、激素调控和抗氧化 需冷量特性ꎬ 相比于落叶猕猴桃需要更少的低温