
Page 206 - 《广西植物》2023年第11期
P. 206
2 1 6 6 广 西 植 物 43 卷
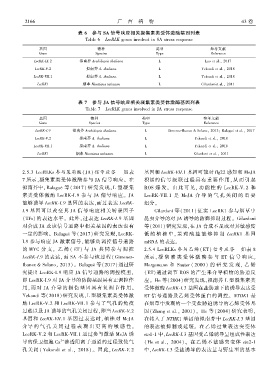
表 6 参与 SA 信号响应相关凝集素类受体激酶基因列表
Table 6 LecRLK genes involved in SA stress response
基因 物种 类型 参考文献
Gene Species Type Reference
LecRK ̄IX.2 拟南芥 Arabidopsis thaliana L Luo et al.ꎬ 2017
LecRK ̄V.2 拟南芥 A. thaliana L Yekondi et al.ꎬ 2018
LecRK ̄VII.1 拟南芥 A. thaliana L Yekondi et al.ꎬ 2018
LecRK1 烟草 Nicotiana tabacum L Gilardoniet al.ꎬ 2011
表 7 参与 JA 信号响应相关凝集素类受体激酶基因列表
Table 7 LecRLK genes involved in JA stress response
基因 物种 类型 参考文献
Gene Species Type Reference
LecRK ̄I.9 拟南芥 Arabidopsis thaliana L Gimenez ̄Ibanez & Solanoꎬ 2013ꎻ Balagué et al.ꎬ 2017
LecRK ̄V.2 拟南芥 A. thaliana L Yekondi et al.ꎬ 2018
LecRK ̄VII.1 拟南芥 A. thaliana L Yekondi et al.ꎬ 2018
LecRK1 烟草 Nicotiana tabacum L Gilardoni et al.ꎬ 2011
2.5.3 LecRLKs 参与茉莉酸( JA) 信号应答 如表 基因和 LecRK ̄VII.1 基因可能对 flg22 感知和 MeJA
7 所示ꎬ凝集素类受体激酶参与 JA 信号响应ꎮ 在 积累的 信 号 级 联 过 程 具 有 重 要 作 用ꎬ 从 而 引 起
拟南芥中ꎬBalagué 等( 2017) 研究发现ꎬL 型凝集 ROS 爆 发ꎮ 由 此 可 见ꎬ 功 能 性 的 LecRK ̄V. 2 和
素类受体激酶 LecRK ̄I.9 参与 JA 信号响应ꎮ JA LecRK ̄VII.1 是 MeJA 介 导 的 气 孔 关 闭 的 重 要
能够诱导 LecRK ̄I.9 基因的表达ꎬ而过表达 LecRK ̄ 组分ꎮ
I.9 基因可以改变其 JA 信号响应相关转录因子 Gilardoni 等(2011) 证实 LecRK1 参与烟草中
(TFs) 的表达水平ꎮ 此外ꎬ过表达 LecRK ̄I.9 基因 昆虫介导的对 JA 诱导的防御抑制过程ꎮ Gilardoni
对合成 JA 及该信号通路中相关基因的表达也有 等(2011)研究发现ꎬ在 JA 合成不足或对其敏感度
一定的影响ꎮ Balagué 等(2017) 研究发现ꎬLecRK ̄ 低 的 植 株 中ꎬ 茉 莉 酸 盐 能 够 抑 制 LecRK1 基 因
I.9 参与响应 JA 激素信号ꎬ能够负调控信号通路 mRNA 的表达ꎮ
的 MYC 分 支ꎮ 乙 烯 ( ET) 与 JA 共 同 参 与 调 控 2.5.4 LecRLKs 参与乙烯( ET) 信号应答 如表 8
LecRK ̄I.9 的表达ꎬ而 SA 不参与该过程( Gimenez ̄ 所示ꎬ 凝 集 素 类 受 体 激 酶 参 与 ET 信 号 响 应ꎮ
Ibanez & Solanoꎬ 2013)ꎮ Balagué 等(2017)通过研 Mergemann 和 Sauter ( 2000 ) 的 研 究 发 现ꎬ 乙 烯
究提出 LecRK ̄I.9 响应 JA 信号通路的调控模型ꎬ (ET)通过调节 ROS 的产生来介导植物的胁迫反
即 LecRK ̄I.9 对 JA 介导的防御基因具有正调控作 应ꎮ He 等(2004)研究发现ꎬ拟南芥 L 型凝集素类
用ꎬ 而 对 JA 介 导 的 损 伤 基 因 具 有 负 调 节 作 用ꎮ 受体激酶 LecRK ̄I.3 基因在盐胁迫下的诱导表达受
Yekondi 等(2018)研究发现ꎬL 型凝集素类受体激 ET 信号通路及乙烯受体蛋白的调控ꎮ NTHK1 是
酶 LecRK ̄V.2 和 LecRK ̄VII.1 参与了气孔的免疫 在烟草中发现的一个受盐胁迫诱导的乙烯受体基
过程以及 JA 诱导的气孔关闭过程ꎬ即当 LecRK ̄V.2 因(Zhang et al.ꎬ 2001)ꎮ He 等(2004) 研究表明ꎬ
基因和 LecRK ̄VII.1 基因过表达时ꎬ植株对 MeJA 在转入了 NTHK1 基因的拟南芥中 LecRK ̄I.3 基因
介导 的 气 孔 关 闭 过 程 表 现 出 更 高 的 敏 感 性ꎮ 的表达被抑制或延缓ꎮ 在乙烯过量表达突变体
LecRK ̄V.2 和 LecRK ̄VII.1 通过参与激活 MeJA 诱 eto1 ̄1 中ꎬLecRK ̄I.3 基因受乙烯诱导呈组成性表达
导的保卫细胞 Ca 渗透阳离子通道的过程致使气 (He et al.ꎬ 2004)ꎮ 在乙烯不敏感突变体 ein2 ̄1
2+
孔关 闭 ( Yekondi et al.ꎬ 2018)ꎮ 因 此ꎬLecRK ̄V. 2 中ꎬLecRK ̄I.3 受盐诱导的表达量与野生型的基本