
Page 203 - 《广西植物》2023年第11期
P. 203
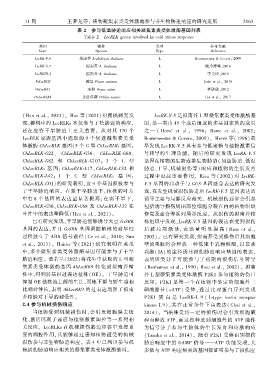
11 期 王梦龙等: 植物凝集素类受体激酶参与非生物胁迫响应的研究进展 2 1 6 3
表 2 参与低温胁迫响应相关凝集素类受体激酶基因列表
Table 2 LecRLK genes involved in cold stress response
基因 物种 类型 参考文献
Gene Species Type Reference
LecRK ̄V.6 拟南芥 Arabidopsis thaliana L Bouwmeester & Goversꎬ 2009
LecRK ̄S.7 拟南芥 A. thaliana L 陆秀涛等ꎬ2016
LecRKIII.2 拟南芥 A. thaliana L 李美玲ꎬ2019
PsLecRLK 豌豆 Pisum sativum L Joshi et al.ꎬ 2010
OsLecRK1 水稻 Oryza sativa L 崔欣欣ꎬ2012
PnLecRLK1 黄丝瓜藓 Pohlia nutans L Liu et al.ꎬ 2017
(Han et al.ꎬ 2021)ꎮ Han 等(2021) 对樱桃研究发 LecRK ̄V.5 是拟南芥 L 型凝集素类受体激酶基
现ꎬ樱桃中的 LecRLKs 不仅参与了盐胁迫的响应ꎬ 因ꎬ是一类由 45 个成员组成的多基因家族的成员
还在应答干旱胁迫上至关重要ꎮ 从对其 170 个 之 一 ( Hervé et al.ꎬ 1996ꎻ Barre et al.ꎬ 2002ꎻ
LecRLK 家族基因中选取的 9 个候选凝集素类受 Bouwmeester & Goversꎬ 2009)ꎮ Hervé 等(1996)最
体激酶 ChLecRLK 基因(5 个 G 型 ChLecRLKs 基因: 早发现 LecRK ̄V.5 具有参与低聚糖与植物激素信
ChLecRLK ̄G22、 ChLecRLK ̄G36、 ChLecRLK ̄G68、 号转导的生理功能ꎮ 随后的研究发现 LecRK ̄V.5
ChLecRLK ̄G82 和 ChLecRLK ̄G107ꎻ 3 个 L 型 基因在植物的生物或非生物胁迫( 如盐胁迫、低温
ChLecRLKs 基 因: ChLecRLK ̄L17、 ChLecRLK ̄L32 和 胁迫、干旱、机械损伤等) 响应和植物的生长发育
ChLecRLK ̄L42ꎻ 1 个 C 型 ChLecRLKs 基 因: 过程中也起重要作用ꎮ Riou 等( 2002) 对 LecRK ̄
ChLecRLK ̄C01)的研究表明ꎬ这 9 个基因积极参与 V.5 基因的启动子与 GUS 基因融合表达的研究发
了干旱胁迫响应ꎮ 在强干旱胁迫下ꎬ植株的叶片 现ꎬ在发生机械损伤胁迫后 LecRK ̄V.5 基因表达的
中有 8 个基因的表达量显著提高ꎻ在弱干旱下ꎬ 诱导主要与局部反应相关ꎮ 机械损伤通常会引起
ChLecRLK ̄G36、ChLecRLK ̄G68 及 ChLecRLK ̄L32 在 包括诱导损伤周围部位细胞分裂在内的植物组织
叶片中的表达量降低(Han et al.ꎬ 2021)ꎮ 修复及愈合等系列局部反应ꎮ 从损伤的拟南芥植
已有研究发现ꎬ干旱胁迫能够诱导大豆 GsSRK 株组织中发现ꎬLecRK ̄V.5 基因的表达在受到损伤
基因的表达ꎬ并且 GsSRK 基因调控植株的耐旱性 后 被 局 部 激 活ꎬ 表 达 量 明 显 提 高 ( Riou et al.ꎬ
过程独立于 ABA 信号途径( Ge et al.ꎬ 2010ꎻ Sun 2002)ꎮ 已有研究发现ꎬ拟南芥受到损伤后其细胞
et al.ꎬ 2013)ꎮ Haider 等(2021) 研究表明在黄瓜 壁的果胶将会释放一种低聚半乳糖醛酸ꎬ以非茉
中ꎬ多个凝集素类受体激酶基因可能参与了干旱 莉酸(JA)的途径诱导损伤胁迫响应基因的表达ꎬ
胁迫响应ꎮ 黄若兰(2022)将花生中获取的 L 型凝 表明该类分子可能参与了初期的损伤信号转导
集素类受体激酶基因 AhLecRK9 转化到拟南芥植 (Benhamou et al.ꎬ 1990ꎻ Riou et al.ꎬ 2002)ꎮ 拟南
株中ꎬ得到转基因过表达植株( OE)ꎮ 干旱胁迫可 芥 L 型凝集素类受体激酶 P2K1 参与植物的伤口
抑制 OE 植株地上部的生长ꎬ而地下部与野生型相 反应ꎮ P2K1 是第一个在植物中鉴定的细胞外三
比相对伸长ꎬ表明 AhLecRK9 的过表达增强了拟南 磷酸腺苷 ( eATP ) 受 体ꎬ通 过 比 对 蛋 白 序 列 发 现
芥植株对干旱的耐受性ꎮ P2K1 蛋 白 是 LecRK ̄I. 9 ( L ̄type lectin receptor
2.4 参与机械损伤响应 kinase I.9)ꎬ其在正常条件下高表达( Choi et al.ꎬ
当植物受到机械损伤时ꎬ会引发细胞膜去极 2014)ꎮ 当植株受到一定的损伤时会引发细胞破
化、激活钙离子通道与植物激素调控等一系列相 碎而释放 ATPꎬ而这些释放到细胞外的 ATP 能作
关反应ꎮ LecRLKs 在机械损伤胁迫应答中发挥重 为信号分子参与生物体的生长发育和应激响应
要的调控作用ꎬ其能够通过感知植物遭受的机械 (Tanaka et al.ꎬ 2014)ꎮ 随着 P2K1 受体识别损伤
损伤参与非生物胁迫响应ꎮ 表 4 中已列出参与机 胁迫响应中的 DAMP 信号———ATP 功能发现ꎬ大
械损伤胁迫响应相关的凝集素类受体激酶基因ꎮ 多数与 ATP 响应相关的基因被证明参与了损伤应