
Page 185 - 《广西植物》2023年第9期
P. 185
9 期 李文杨等: 不同光质与光周期对粗肋草生长、叶片色素和颜色的影响 1 7 3 3
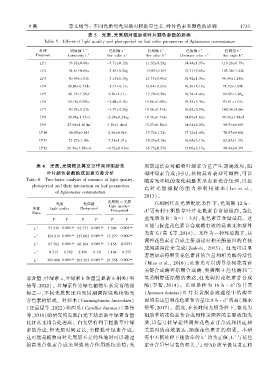
表 5 光质、光周期对粗肋草叶片颜色参数的影响
Table 5 Effects of light quality and photoperiod on leaf color parameters of Aglaonema commutatum
处理 明度值 L ∗ 色相值 a ∗ 色相值 b ∗ 色度值 c ∗ 色调角 h ∗
Treatment Luminosity L ∗ Hue value a ∗ Hue value b ∗ Chromatic value c ∗ Hue angle h ∗
LP1 35.62±0.80a -5.71±0.12h 11.52±0.29g 14.44±1.55a 110.20±0.79a
LP2 34.61±0.60a -3.83±0.24g 13.03±0.39f 15.71±5.69a 103.36±1.42b
LP3 30.95±1.01b -3.15±0.19g 13.77±0.45ef 18.42±1.36a 96.59±1.25de
LP4 28.06±0.73de -1.71±0.11e 13.84±0.33ef 16.36±6.19a 74.72±1.09h
LP5 26.15±1.56ef 2.20±1.11c 17.79±0.50a 18.78±1.49a 84.69±1.26g
LP6 30.16±0.99bc -2.04±0.18e 15.86±0.69bc 19.55±5.36a 93.81±1.01e
LP7 30.92±0.21b -3.97±0.20g 14.36±0.51de 16.62±0.94a 100.36±4.66c
LP8 28.86±1.53cd -2.28±0.21eg 14.36±0.33de 14.07±1.62a 98.92±1.88cd
LP9 27.98±1.61def 3.29±1.48cd 15.27±0.50cd 14.94±0.25a 90.51±0.99f
LP10 26.05±0.84f 4.26±0.94b 15.72±1.21c 17.22±1.45a 70.87±0.60i
LP11 21.27±1.10h 7.51±1.31a 18.35±0.18a 16.64±5.15a 62.83±1.33j
LP12 29.34±1.26bcd -0.52±0.67de 16.75±0.55b 13.09±2.19a 90.04±0.39f
表 6 光质、光周期及其交互作用对粗肋草 周期过长会对植物叶绿素含量产生逆境效应ꎬ阻
叶片颜色参数的双因素方差分析 碍叶绿素合成ꎻ同时ꎬ植物具有表型可塑性ꎬ可以
Table 6 Two ̄factor analysis of variance of light qualityꎬ 随着光环境的变化调整其形态和光合生理ꎬ以提
photoperiod and their interaction on leaf parameters
高对 光 能 捕 捉 的 能 力 和 利 用 效 率 ( Lei et al.ꎬ
of Aglaonema commutatum
2013)ꎮ
光周期 × 光质
光质 光周期 在相同红蓝光质配比条件下ꎬ光周期 12 h
名称 Light quality Photoperiod Light quality×  ̄1
Photoperiod d 更有利于粗肋草叶片花色素苷含量提高ꎬ当红
Name
蓝光质为 R ∶ B = 1 ∶ 3 时ꎬ花色素苷含量最高ꎮ 这
F P F P F P
可能与促进花色素苷合成最有效的是蓝光和紫外
L ∗ 57.936 0.000 92.733 0.000 4.506 0.005
光有关(高飞等ꎬ2014)ꎮ 光作为一种环境因子ꎬ其
a ∗ 124.810 0.000 227.002 0.000 15.279 0.000
调控花色素苷合成主要通过对相关酶基因的直接
b ∗ 67.762 0.000 66.204 0.000 3.156 0.025
或间接调控来完成( Jaakolaꎬ 2013)ꎮ 红光可以显
c ∗ 0.515 0.762 2.196 0.151 1.164 0.355
著增加草莓果实花色素苷的含量和相关酶的活性
h ∗ 369.008 0.000 204.563 0.000 24.358 0.000
(Miao et al.ꎬ 2016)ꎻ蓝紫光可以诱导茶树花青苷
关键合成酶查尔酮合成酶、类黄酮 ̄3 ̄羟化酶和二
总含量、叶绿素 a、叶绿素 b 含量呈显著正相关( 项 氢黄酮醇还原酶的表达ꎬ红光调控花色素苷合成
倩等ꎬ2022)ꎮ 叶绿素作为绿色植物生长发育的指 酶(李 智ꎬ2014)ꎮ 光 照 条 件 为 16 h d 的 甘 薯
 ̄1
标之一ꎬ不同光质配比和光周期调控绿色植物光 (Ipomoea batatas) 叶片类黄酮合成途径中结构基
 ̄1
合色素的形成ꎮ 对杉木( Cunninghamia lanceolata) 因的表达量和花色素苷含量比 8 hd 的高( 魏永
(汪星星等ꎬ2022) 和西瓜( Citrullus lanatus) ( 秦伟 赞等ꎬ2017)ꎮ 因此ꎬ在长时间光照条件下ꎬ蓝光为
等ꎬ2018)的研究均发现白光下幼苗的叶绿素含量 粗肋草的花色素苷合成和转录调控的主要效能光
比红蓝光组合处理高ꎮ 白光更有利于粗肋草叶绿 质ꎬ以信号转导途径调控花色素苷合成和转运相
素的合成ꎬ但光照时间过长ꎬ会降低叶绿素合成ꎮ 关基因的高效表达ꎬ加强花色素苷的积累ꎮ 本研
这可能是植物面对光周期不足的环境时可以通过 究中不同处理下粗肋草的 L 均为正值ꎬL 与花色
∗
∗
提高光合色素合成来增强光合作用抵抗逆境ꎻ光 素苷含量呈显著负相关ꎬ与 SPAD 值呈极显著正相