
Page 10 - 《广西植物》2024年第12期
P. 10
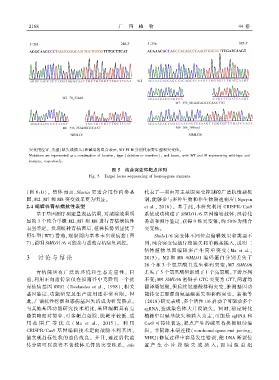
2 1 6 8 广 西 植 物 44 卷
突变用位置、类型(缺失或插入)和碱基的组合表示ꎬWT 和 M 分别代表野生型和突变体ꎮ
Mutations are represented as a combination of locationꎬ type (deletion or insertion)ꎬ and basesꎬ with WT and M representing wild ̄type and
mutantsꎬ respectively.
图 5 纯合突变体靶点序列
Fig. 5 Target locus sequencing of homozygous mutants
(图 6:D)ꎮ 整体而言ꎬSlactin 更适合用作内参基 代表了一类由寄主基因突变控制的广谱抗性新机
因ꎬM2、M7 和 M8 突变效果更为明显ꎮ 制ꎬ能够参与多种生物和非生物胁迫响应( Nguyen
2.4 编辑株青枯病抗性表型 et al.ꎬ 2016)ꎮ 基于此ꎬ本研究利用 CRISPR / Cas9
基于基因测序和定量表达结果ꎬ对敲除效果明 系统成功构建了 SlMLO1 / 6 基因编辑载体ꎬ经转化
显的 3 个纯合单株 M2、M7 和 M8 进行青枯病抗性 番茄和测序鉴定ꎬ获得 9 株突变体ꎬ约 50%为纯合
表型鉴定ꎮ 发现接种青枯菌后ꎬ植株长势明显优于 突变株ꎮ
野生型( WT) 番茄ꎬ观察期内基本未出现病症( 图 Slmlo1 / 6 突变体不同位点编辑效果和类型不
7)ꎬ说明 SlMLO1 / 6 可能参与番茄青枯病负调控ꎮ 同ꎬ纯合突变包括片段缺失和单碱基插入ꎬ说明二
倍体植 物 基 因 编 辑 多 产 生 简 单 突 变 ( Ma et al.ꎬ
3 讨论与结论 2015)ꎮ M2 和 M8 SlMLO1 编码蛋白分别丢失了
20 个和 3 个氨基酸且发生移码突变ꎻM7 SlMLO6
青枯 菌 具 有 广 泛 的 环 境 和 生 态 适 应 性ꎮ 目 丢失了 5 个氨基酸但新增了 1 个亮氨酸ꎬ下游序列
前ꎬ利用正向遗传学仅在拟南芥中克隆到一个抗 不变ꎻM9 SlMLO6 密码子 GTG 突变为 GTTꎬ两者均
青枯病基因 RRS1 ( Deslandes et al.ꎬ 1998)ꎬ相关 翻译缬氨酸ꎬ但后续氨基酸移码突变ꎬ推测基因功
基因鉴定、功能研究及生产应用还非常有限ꎮ 因 能转变主要源自氨基酸丢失和移码突变ꎮ 蒲艳等
此ꎬ广谱抗性挖掘和感病基因失活成为研究热点ꎮ (2018)研究表明ꎬ多个活性 U6 启动子可驱动多个
与其他基因功能研究技术相比ꎬ基因编辑具有克 sgRNAꎬ造成染色体大片段缺失ꎮ 同时ꎬ稳定转化
隆策略相对简单、可多靶点敲除、脱靶率较低、适 植株中以碱基缺失和插入为主ꎬ可能是 sgRNA 和
用 范 围 广 等 优 点 ( Ma et al.ꎬ 2015 )ꎮ 利 用 Cas9 可持续表达ꎬ靶点产生的碱基替换被继续编
CRISPR / Cas9 基因编辑技术定向敲除不利基因ꎬ 辑ꎮ 非同源末端连接( non ̄homologous end joiningꎬ
能实现目标性状的遗传改良ꎮ 并且ꎬ通过后代遗 NHEJ)修复过程中容易发生错误ꎬ使 DNA 断裂位
传分离可以获得不含载体元件的突变株系ꎮ mlo 置 产 生 小 片 段 缺 失 或 插 入ꎻ 而 同 源 重 组