
Page 176 - 《广西植物》2024年第5期
P. 176
9 6 4 广 西 植 物 44 卷
吸光值ꎬ按标准曲线计算种子中的蛋白含量ꎮ 热 周含水量显著降低(P<0.05)ꎬ下降速度迅猛ꎬ平均
稳定蛋白则按 50 μL 样品加同等的提取液稀释ꎬ 每周降低 2.8%ꎬ从 41 周开始维持在相对稳定的水
再加入 0.5 mL 的考马斯亮蓝染液混匀ꎬ静置 5 ~ 20 平(约 42%)ꎮ 种子干鲜重比在整个发育过程中呈
minꎬ用酶标仪在 595 nm 处测定 OD 值ꎬ根据标准 现出从上升到稳定的变化ꎬ花后 23 ~ 27 周种子的
曲线计算蛋白含量ꎮ 干鲜重比保持在 17%左右ꎬ此后至 41 周呈逐渐上
1.2.4 不同发育时期种子胚轴细胞超显微结构观 升的 趋 势ꎬ 到 花 后 41 周 时 种 子 的 干 鲜 重 比 达
察 种子最重要的使命是萌发和发育成幼苗ꎬ而 57%ꎬ增加了近 39%ꎬ此后趋于稳定ꎮ 以上结果说
胚轴是种子最重要组成部分ꎬ是种子能否发育成 明ꎬ‘曼赛龙柚’种子在花后 41 周前后达到生理性
为正常幼苗的关键部位ꎮ 种子在受到逆境胁迫 成熟ꎬ并且在此之后种子的含水量趋于稳定ꎬ稳定
时ꎬ胚轴受到伤害最容易导致丧失活力( Priestleyꎬ 在 42%左右ꎮ 种子成熟时含水量依然很高ꎬ说明
1986ꎻ 程红焱等ꎬ1991)ꎮ 前人关于种子超显微结 该种子没有成熟脱水过程ꎮ
构的研究也经常是以胚(对胚比较小的种子) 或胚
轴( 对胚比较大的种子) 开展的ꎬ如玉米( Wen et
al.ꎬ 2009) 和蒲葵( Wenꎬ2011)ꎮ 为此ꎬ从上述不
同发育时期的‘曼赛龙柚’ 种子中取 5 粒种子作为
样品ꎬ用解剖刀和镊子去除种皮并切下种子包含
胚轴的尖端ꎮ 取 3 mL 电镜固定液对样品进行固
定并保存在 4 ℃ 下备用ꎮ 在电镜观察前需要先制
作超薄切片ꎬ切取约 1 mm 的胚根组织ꎬ用 3.5%戊
二醛和 1%的锇酸固定ꎬ用丙酮逐级脱水后将材料
放进环氧树脂中包埋 1 周ꎮ 材料处理好后先切成
半薄切片ꎬ于光学显微镜下筛选出完好的样品制
所有数据为 10 个或 8 个重复ꎬ以平均值±标准误表示ꎮ
作超薄切片( 王若兰等ꎬ2014)ꎮ 将制作好的超薄
All data are expressed as x±s x of 10 or 8 replicates.
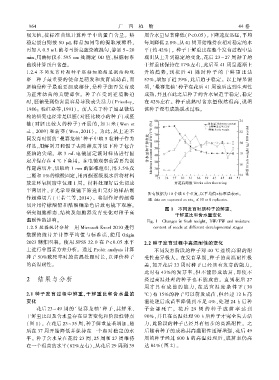
切片用柠檬酸铅和醋酸铀染色后放电镜下观察ꎮ 图 1 不同发育时期种子的鲜重、
研究细胞形态、结构及细胞器发育变化对种子高
干鲜重比和含水量变化
温耐性的影响ꎮ Fig. 1 Changes in fresh weightꎬ DW/ FW and moisture
1.2.5 数据统计分析 用 Microsoft Excel 2010 进行 content of seeds at different developmental stages
数据的统计并计算平均值与标准误ꎬ使用 Origin
2023 制作图表ꎮ 使用 SPSS 22.0 在 P<0.05 水平 2.2 种子发育过程中高温耐性的变化
上进行单因素方差分析ꎮ 通过 Probit analysis 计算 不同发育阶段的种子对 40 ℃ 连续高温的耐
种子 50%致死率时的高温处理时长ꎬ以评价种子 受性差异极大ꎮ 在发育早期ꎬ种子的高温耐性极
的高温耐性ꎮ 差ꎬ如开花后 23 周时种子已经具有发芽的能力ꎬ
此时有 43%的发芽率ꎬ但不能形成幼苗ꎬ即使不
2 结果与分析 经过高温处理的种子也不能成苗ꎮ 直到花后 27
周才 具 有 成 苗 的 能 力ꎬ 在 适 宜 温 度 条 件 下 ( 30
2.1 种子发育过程中鲜重、干鲜重比和含水量的 ℃ ) 有 15%的种子可以萌发成苗ꎬ但经过 12 h 高
变化 温处理后成苗率降低到不足 4%ꎬ处理 24 h 后种
花后 23 ~ 49 周的‘ 曼赛龙柚’ 种子ꎬ其鲜重、 子全 部 死 亡ꎮ 花 后 29 周 的 种 子 成 苗 率 达 到
干鲜重比以及含水量存在显著变化和阶段性特点 90%ꎬ并且在高温处理 90 h 后种子才完全失去活
(图 1)ꎮ 在花后 23 ~ 35 周ꎬ种子鲜重显著增加ꎬ随 力ꎬ此阶段的种子已经具有初步的高温耐性ꎮ 之
后在 37 周开始降低并保持在一个相对稳定的水 后随着种子的成熟其高温耐性逐渐增强ꎬ花后 49
平ꎮ 种子含水量在花后 23 周、25 周和 27 周维持 周的种子经过 600 h 的高温处理后ꎬ成苗率仍高
在一个很高的水平(82%左右)ꎬ从花后 29 周到 39 达 84%( 图 2) ꎮ