
Page 106 - 《广西植物》2025年第1期
P. 106
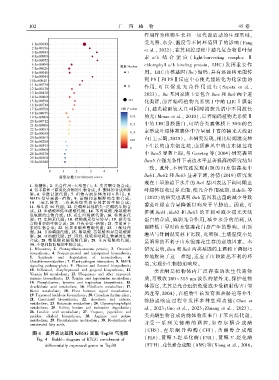
1 0 2 广 西 植 物 45 卷
作用作为植物生长和一切代谢活动的生理基础ꎬ
受光照、水分、温度等不同环境因子的影响( Fang
et al.ꎬ 2022)ꎬ在其调控过程中捕光复合物Ⅱ叶绿
素 a / b 结 合 蛋 白 ( light ̄harvesting complex Ⅱ
chlorophyll a / b binding proteinꎬ LHC) 发挥重要作
用ꎮ LHC 由核基因( lhc) 编码ꎬ具有迅速将光能传
到 PSⅠ和 PSⅡ反应中心使光能转化为化学能的
作用ꎬ 可 以 促 进 光 合 作 用 进 行 ( Sapeta et al.ꎬ
2023)ꎮ lhc 基因家族主要包含 lhca 和 lhcb 两个进
化类群ꎬ前者编码植物光系统 I 中的 LHCⅠ膜蛋
白ꎬ捕光截面较大且可同时捕获光谱中不同波长
的光(Mozzo et al.ꎬ 2010)ꎬ后者编码植物光系统Ⅱ
中的 LHCⅡ膜蛋白ꎬ可结合类囊体膜上 50%的色
素形成叶绿体类囊体中含量最丰富的捕光天线蛋
白(王云鹤ꎬ2020)ꎮ 本研究发现ꎬ相比针阔混交林
下生长的山东银莲花ꎬ山顶灌丛中的山东银莲花
中 lhca5 显著上调ꎬ与 Ganeteg 等(2004) 研究表明
lhca5 在强光条件下表达水平显著提高的研究结果
一致ꎮ 此外ꎬ本研究还发现山顶的山东银莲花中
lhcb1、lhcb2 和 lhcb3 显著下调ꎬ蒋倩(2019) 研究发
现在干旱胁迫下水芹的 lhcb 基因表达下调可阻止
1. 核糖体ꎻ 2. 光合作用-天线蛋白ꎻ 3. 类黄酮生物合成ꎻ
4. 倍半萜和三萜类化合物的生物合成ꎻ 5. 酮体的合成和降 叶绿体接收过多光能ꎬ使光合作用减弱ꎬRahele 等
解ꎻ 6. 谷胱甘肽代谢ꎻ 7. 植物与病原体的相互作用ꎻ 8.
MAPK 信号 通 路 - 植 物ꎻ 9. 黄 酮 和 黄 酮 醇 的 生 物 合 成ꎻ (2022)的研究也表明 lhcb 基因表达量的减少导致
10. 二苯 乙 烯 类、 二 芳 基 庚 烷 类 和 姜 辣 素 的 生 物 合 成ꎻ 番茄叶绿素含量降低以响应干旱胁迫ꎮ 因此ꎬ可
11. 维生素 B6 代谢ꎻ 12. 泛醌和其他萜类-泛醌的生物合
成ꎻ 13. 牛磺酸和低牛磺酸代谢ꎻ 14. 苯丙氨酸、酪氨酸和 推测 lhcb1、lhcb2 和 lhcb3 的下调可减少捕光天线
色氨酸的生物合成ꎻ 15. 花生四烯酸代谢ꎻ 16. 核黄素代
谢ꎻ 17. 生物素代谢ꎻ 18. 植物激素信号转导ꎻ 19. 萜类化 蛋白的合成ꎬ减弱光合作用ꎬ减少水分的消耗ꎬ从
合物骨架的生物合成ꎻ 20. 昼夜节律-植物ꎻ 21. 类胡萝卜
素的生物合成ꎻ 22. 抗坏血酸和醛酸代谢ꎻ 23. 丁酸盐代 而降低干旱对山东银莲花自身产生的影响ꎮ 山顶
谢ꎻ 24. 甘油磷脂代谢ꎻ 25. 缬氨酸、亮氨酸和异亮氨酸降
解ꎻ 26. 亚油酸代谢ꎻ 27. 丙烷、哌啶和吡啶生物碱的生物 灌丛与针阔混交林下比较ꎬ光照强、土壤湿度小是
合成ꎻ 28. 精氨酸和脯氨酸代谢ꎻ 29. 苯 丙 氨 酸 的 代 谢ꎻ 最明显的不利于山东银莲花生存的逆境因素ꎮ 本
30. 不饱和脂肪酸的生物合成ꎮ
研究表明ꎬlhca 和 lhcb 两类基因的上调和下调很巧
1. Ribosomeꎻ 2. Photosynthesis ̄antenna proteinꎻ 3. Flavonoid
biosynthesisꎻ 4. Sesquiterpenoid and triterpenoid biosynthesisꎻ
5. Synthesis and degradation of ketonebodiesꎻ 6. 妙地解决了这一难题ꎬ适应了山顶灌丛不利的环
Glutathionemetabolismꎻ 7. Plant ̄pathogen interactionꎻ 8. MAPK
境ꎬ实现非生物胁迫响应ꎮ
signaling pathway ̄plantꎻ 9. Flavone and flavonol biosynthesisꎻ
10. Stilbenoidꎬ diarylheptanoid and gingerol biosynthesisꎻ 11. 类黄酮是植物体内广泛 存 在 的 次 生 代 谢 物
Vitamin B6 metabolismꎻ 12. Ubiquinone and other terpenoid ̄
quinone biosynthesisꎻ 13. Taurine and hypotaurine metabolismꎻ 质ꎬ可吸收 280 ~ 315 nm 波长的紫外光ꎬ保护植物
14. Phenylalanineꎬ tyrosine and tryptophan biosynthesisꎻ 15.
Arachidonic acid metabolismꎻ 16. Riboflavin metabolismꎻ 17. 体器官ꎬ尤其是光合组织免受或少受辐射伤害( 邹
Biotin metabolismꎻ 18. Plant hormone signal transductionꎻ 凤莲等ꎬ2004)ꎬ在植物生长发育和热胁迫等非生
19. Terpenoid backbone biosynthesisꎻ 20. Circadian rhythm ̄plantꎻ
21. Carotenoid biosynthesisꎻ 22. Ascorbate and aldarate 物胁迫 响 应 过 程 中 发 挥 多 种 生 理 功 能 ( Chen et
metabolismꎻ 23. Butanoate metabolismꎻ 24. Glycerophospholipid
metabolismꎻ 25. Valineꎬ leucine and isoleucine degradationꎻ al.ꎬ 2023ꎻGuo et al.ꎬ 2023ꎻZhuang et al.ꎬ 2023)ꎮ
26. Linoleic acid metabolismꎻ 27. Tropaneꎬ piperidine and
pyridine alkaloid biosynthesisꎻ 28. Arginine and proline 类黄酮生物合成的前体物质来自于苯丙烷代谢ꎬ
metabolismꎻ 29. Phenylalanine metabolismꎻ 30. Biosynthesis of 并受 一 系 列 关 键 酶 的 调 控ꎬ 如 查 尔 酮 合 成 酶
unsaturated fatty acids.
(CHS)、 查 尔 酮 异 构 酶 ( CHI)、 黄 酮 醇 合 成 酶
图 4 差异表达基因 KEGG 富集 Top30 气泡图
(FLS)、黄酮 3 ̄羟基化酶( F3H)、黄酮 3′ ̄羟化酶
Fig. 4 Bubble diagram of KEGG enrichment of
differentially expressed genes in Top30 (F3′H)、花色素合成酶 (ANS)等(Xiong et al.ꎬ 2016ꎻ