
Page 152 - 《广西植物》2025年第11期
P. 152
2 0 9 6 广 西 植 物 45 卷
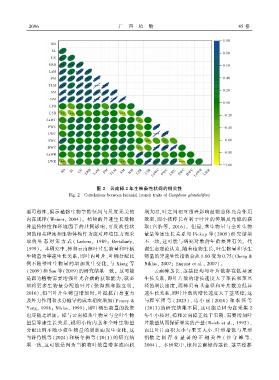
图 2 云南樟 2 年生枝条性状间的相关性
Fig. 2 Correlations between biennial branch traits of Camphora glandulifera
型可塑性ꎬ揭示植物生物学特征间与尺度无关的 增大时ꎬ叶之间相互重叠影响植物总体光合作用
内在规律( Weinerꎬ 2004)ꎮ 植物的异速生长受物 效率ꎬ而小枝伸长有利于叶片的伸展及光能的获
种遗传特性和环境因子的共同影响ꎬ可反映性状 取(宫浩等ꎬ 2016)ꎮ 但是ꎬ茎生物量与全叶生物
间的相关程度和生物体构件为应对环境压力而采 量呈等速生长关系与 Pickup 等(2005) 研究结果
取的 生 态 对 策 方 式 ( Larberaꎬ 1989ꎻ Bertalanfyꎬ 不一致ꎬ这可能与研究对象的年龄差异有关ꎮ 代
1999)ꎮ 本研究中ꎬ樟和云南樟叶片生物量和叶柄 谢生态理论认为ꎬ随着植物生长ꎬ叶生物量和茎生
生物量为等速生长关系ꎬ即叶内叶片、叶柄分配比 物量的异速生长指数会从 1.00 变为 0.75(Cheng &
例不随整叶生物量的增加发生变化ꎬ与 Xiang 等 Niklasꎬ 2007ꎻ Enquist et al.ꎬ 2007)ꎮ
(2009)和 Sun 等(2019)的研究结果一致ꎮ 这可能 云南樟茎长、茎基径均与叶片数存在低异速
是因为植物需要增强叶光合碳的获取能力ꎬ就必 生长关系ꎬ即叶片数的增长速度大于茎长和茎基
须将更多生物量分配给叶片( 张海燕和陈立明ꎬ 径的增长速度ꎬ而樟只有茎基径和叶片数为低异
2016)ꎬ但当叶片生物量增加时ꎬ叶抵抗自身重力 速生长关系ꎬ即叶片数的增长速度大于茎基径ꎬ这
及外力作用和水分输导的成本相应增加( Pearcy & 与翟军 团 等 ( 2023 )、 马 小 丽 ( 2014 ) 和 李 钰 等
Yangꎬ 1998ꎻ Niklasꎬ 1999)ꎬ即叶柄生物量的投资 (2013)的研究结果不同ꎬ这可能是因为在采集 2
也应随之增加ꎮ 樟与云南樟茎生物量与全叶生物 年生小枝时ꎬ樟和云南樟正处于果期ꎬ需要增加叶
量呈等速生长关系ꎬ说明小枝内茎和全叶生物量 片数量从而保证果实的产量( Reich et al.ꎬ 1992)ꎬ
分配比例不随小枝生物量的增加而发生变化ꎬ这 而且叶片面积大小与果实大小、叶形指数与果形
与许竹锐等(2024) 和杨冬梅等(2011) 的研究结 指数 之 间 存 在 显 著 的 正 相 关 性 ( 沙 守 峰 等ꎬ
果一致ꎬ这可能是因为当植物叶数量增多或面积 2004) ꎮ 本研究中ꎬ樟和云南樟的茎长、茎基径都