
Page 171 - 《广西植物》2026年第1期
P. 171
1 期 索南邓登等: 濒危药用植物掌裂兰在青藏高原东源地带种群结构及群落特征分析 1 6 7
0.05)ꎬ无呈正相关物种ꎬ与 20 种物种相关关系不显
著(P>0.05) (表 5) ꎮ
2.3.3 物种多 样 性 变 化 利 用 香 农 多 样 性 指 数
( Shannon ̄Wiener indexꎬ H′) 对 3 个 地 点 的 物 种
多样性进行分析( 表 6) ꎬ结果显示海晏掌裂兰群
落物种多样性高于同德和泽库ꎮ 从辛普森优势
度指数( Simpson’ s dominance indexꎬ D) 分析结果
来看( 表 7) ꎬ以泽库样地最高ꎬ同德样地居中ꎬ海
晏样地最低ꎮ β 多样性指数方面ꎬ泽库最 低ꎬ海
晏最高ꎬ表明 β 多样性指数与总物种数、物种丰
富度呈现协同变化格局ꎮ 海晏样地和同德样地
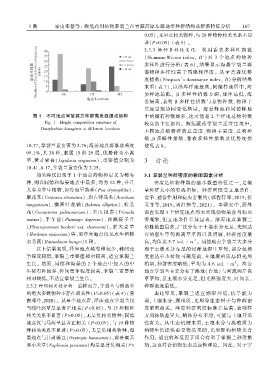
图 1 不同地点间掌裂兰种群高度级组成结构 中的稀有种数最多ꎬ这可能是 2 个样地总物种数
Fig. 1 Height composition structure of
较高的主要原因ꎮ 濒危藏药掌裂兰适宜生境中ꎬ
Dactylorhiza hatagirea at different locations
不同地点植物群落总盖度、物种丰富度、总物种
数、β 多样性指数、香农多样性指数及优势度指
10.77ꎬ掌裂兰重要值为 2.76ꎻ海晏地点群落总盖度 数见表 8ꎮ
99.1%ꎬ共 38 种ꎬ隶属 15 科 29 属ꎬ优势种为小嵩
草、黄帚橐吾( Ligularia virgaurea)ꎬ重要值分别为 3 讨论
18.41、8.17ꎬ掌裂兰重要值为 2.25ꎮ
如果将仅出现于 1 个地点的物种定义为稀有 3.1 掌裂兰种群密度的影响因素分析
种ꎬ则以同德和海晏地点中最多ꎬ均为 12 种ꎬ并且 密度是植物种群的基本数量特征之一ꎬ是衡
大多为中生植物ꎬ如冷地早熟禾( Poa crymophila)、 量种群大小的有效指标ꎮ 种群密度受立地条件、
麻花艽(Gentiana straminea)、唐古特乌头(Aconitum 竞争、捕食作用和病虫害制约( 郭程轩等ꎬ2019ꎻ张
tanguticum)、椭圆叶花锚( Halenia elliptica)、喉 毛 灵菲等ꎬ2019ꎻ刘红梅等ꎬ2021)ꎮ 本研究中ꎬ野外
花( Comastoma pulmonarium)、 天 山 报 春 ( Primula 调查发现 3 个研究地点均未出现动物采食与病虫
nutans)、平车前( Plantago depressa)、西 藏 棱 子 芹 害现象ꎬ但立地条件差异显著ꎮ 泽库地点掌裂兰
(Pleurospermum hookeri var. thomsonii)、 紫 大 麦 草 植株数量最多ꎬ广泛分布于土壤水分充足、光照适
(Hordeum violaceum)等ꎬ而泽库地点仅见水生植物 宜的湿生草甸和嵩草草甸以及阴坡ꎬ种群密度最
 ̄2
水毛茛(Batrachium bungei)1 种ꎮ 高ꎬ均值达 5.7 ind.m ꎮ 同德地点掌裂兰大多分
以上结果表明ꎬ泽库地点稀有种较少ꎬ种间竞 布于土壤水分充足的河滩地湿生草甸ꎬ部分植株
争程度较低ꎬ掌裂兰重要值相对较高ꎬ适宜掌裂兰 受密丛草本植物马蔺庇荫ꎬ土壤腐殖质厚但光照
生长ꎮ 然而ꎬ同德和海晏的 2 个地点中侵入的中 稍弱ꎬ种群密度略低ꎬ平均为 4.6 ind.m ꎮ 海晏
 ̄2
生稀有种较多ꎬ种间竞争程度较高ꎬ掌裂兰重要值 地点掌裂兰主要分布于滩地(台地) 与河流两岸嵩
相对较低ꎬ不适宜掌裂兰生长ꎮ 草草甸ꎬ虽土壤水分充足ꎬ但光照强度大、时间长ꎬ
2.3.2 种间相关性分析 总体而言ꎬ掌裂兰与群落中 种群密度最低ꎮ
的绝大多数物种不存在相关性(P>0.05)(表 4)(索 由此可 见ꎬ掌 裂 兰 适 宜 潮 湿 环 境ꎬ抗 旱 能 力
南邓登ꎬ2020)ꎮ 从单个地点看ꎬ泽库地点掌裂兰仅 弱ꎬ土壤水分、腐殖质、光照等生态因子与种群密
与线叶嵩草呈显著正相关(P<0.05)ꎬ与 15 种物种 度紧密相关ꎮ 泽库种群密度标准差最高ꎬ表明样
相关关系不显著(P>0.05)ꎬ无呈负相关物种ꎻ同德 方间株数差异大ꎬ植株分布不均ꎬ可能与土壤异质
地点仅与马蔺呈显著正相关 (P<0.05)ꎬ与 19 种物 性有关ꎮ 从生态机理来看ꎬ土壤水分与腐殖质为
种相关关系不显著(P>0.05)ꎬ无呈负相关物种ꎻ海 植物生长提供必要物质基础ꎬ光照影响植物光合
晏地点与甘肃棘豆(Oxytropis kansuensis)、黄帚橐吾 作用ꎬ 适宜的环境因子组合有利于掌裂兰种群繁
和小米草(Euphrasia pectinata)均呈显著负相关(P< 衍ꎬ这也符合植物生态适应性理论ꎮ 因此ꎬ 对于掌