
Page 162 - 《广西植物》2026年第3期
P. 162
5 3 8 广 西 植 物 46 卷
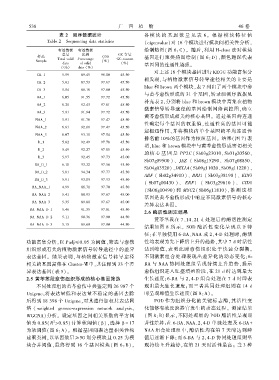
表 2 测序数据统计 各模 块 的 基 因 数 量 见 表 6ꎮ 根 据 模 块 特 征 值
Table 2 Sequencing data statistics (eigenvalue)对 16 个模块进行模块间相关性分析ꎬ
有效数据 有效数据 绘制热图(图 6:C)ꎮ 随后ꎬ利用 H ̄clust 法对模块
总量 比例 GC 含量
样品 Q30 基因进行聚类热图绘制( 图 6:D)ꎬ颜色越深代表
Total valid Percentage GC content
Sample (%)
data of valid (%)
基因间的连通性越强ꎮ
(Gb) data (%)
对上述 16 个模块基因进行 KEGG 功能富集分
CK_1 5.59 89.45 98.00 45.50
析发现ꎬ与植物激素信号转导途径相关的主要是
CK_2 5.61 87.53 97.67 45.50
blue 和 brown 两个模块ꎬ表 7 列出了两个模块中参
CK_3 5.54 88.16 97.80 45.50
与黄芩愈伤形成的 31 个基因ꎬ转录组测序数据见
BA_1 6.05 91.55 97.72 45.50
补充表 2ꎬ分别将 blue 和 brown 模块中富集在植物
BA_2 6.20 92.43 97.61 45.50
激素信号转导途径的基因绘制网络调控图ꎬ确立
BA_3 5.81 91.94 97.72 45.50
黄芩愈伤形成相关的核心基因ꎮ 通过基因的连通
NAA_1 5.91 91.76 97.47 45.50
性确定每个基因的权重值ꎬ连通性高的基因可能
NAA_2 6.61 92.01 97.47 45.50
起枢纽作用ꎬ并将模块内单个基因的平均连通性
NAA_3 6.07 93.10 97.54 45.50
排名前 10%的基因作为枢纽基因ꎮ 结果( 图 7) 表
D_1 5.62 92.49 97.78 45.50
明ꎬblue 和 brown 模块中与黄芩愈伤形成密切相关
D_2 5.49 92.27 97.85 45.50
的核 心 基 因 是 PP2C ( Sb03g23010、 Sb01g20560、
D_3 5.97 92.45 97.73 45.00
Sb07g09500 )、 JAZ ( Sb08g15290、 Sb07g08850、
BA_D_1 6.10 93.32 97.56 45.50
Sb04g03520)、DELLA( Sb09g11020、Sb09g11220)、
BA_D_2 5.81 94.34 97.77 45.50
ABF ( Sb02g34910 )、 BRI1 ( Sb03g38190 )、 EIN3
BA_D_3 5.91 93.09 97.93 45.50
( Sb07g00430 )、 ERF1 ( Sb03g29610 )、 GID1
BA_NAA_1 4.99 88.71 97.70 45.50
(Sb06g10490) 和 MYC2 ( Sb06g11810)ꎬ推 测 这 些
BA_NAA_2 5.41 88.93 97.67 45.00
基因是黄芩愈伤形成中响应不同激素信号的核心
BA_NAA_3 5.35 89.80 97.67 45.00
差异表达基因ꎮ
BA_NAA_D_1 5.46 91.35 97.81 45.50
2.6 酶活性测定结果
BA_NAA_D_2 5.11 90.36 97.90 44.50
黄芩茎段在 7、14、21 d 处理后的酶活性测定
BA_NAA_D_3 5.15 89.69 97.90 44.50
结果如图 8 所 示ꎮ SOD 酶 活 性 变 化 呈 现 以 下 特
征:在单独使用 6 ̄BA、NAA 或 2ꎬ4 ̄D 处理时ꎬ酶活
功能富集分析ꎬ以 Padj≤0.05 为阈值ꎬ筛选与愈伤 性均表现为先下降后上升的趋势ꎬ其中 7 d 时活性
组织形成有关的植物激素信号转导途径中的差异 达到峰值ꎬ表明此时愈伤组织处于快速分裂期ꎮ
表达基因ꎮ 结果表明ꎬ与植物激素信号转导途径 不同激素组合处理表现出差异化的动态变化:6 ̄
相关的基因富集在 Cluster ̄Ⅶ中ꎬ共注释到 33 个差 BA 与 NAA 协同处理组呈现持续上升趋势ꎬ提示
异表达基因(表 5)ꎮ 愈伤组织进入旺盛增殖阶段ꎬ在 21 d 时达到最大
2.5 黄芩茎段愈伤组织形成的核心基因筛选 生长速度ꎻ6 ̄BA 与 2ꎬ4 ̄D 组合处理在 7 d 时即表
不同处理组的黄芩愈伤中共鉴定到 26 987 个 现出最大生长速度ꎻ而三者共同处理组则在 14 d
Unigeneꎬ将表达量低和表达量不稳定的基因去除 时呈现峰值生长速度(图 8:A)ꎮ
后得到 18 398 个 Unigeneꎬ对其进行加权共表达网 POD 作为组织分化的关键标志酶ꎬ其活性变
络 ( weighted geneco ̄expression network analysisꎬ 化能够有效反映器官发生的动态过程ꎮ 测定结果
WGCNA)分析ꎮ 设定基因之间相关系数的平方阈 (图 8:B)显示ꎬ不同处理组的 POD 酶活性呈现显
2
值为 0.85(R >0.85) 计算软阈值( β)ꎬ选择 β = 17 著性差异:在 6 ̄BA、NAA、2ꎬ4 ̄D 单独处理及 6 ̄BA+
为软阈值(图 6:A)ꎮ 根据基因间表达量相关性构 NAA 组合处理组中ꎬ酶活性均在第 7 天时达到峰
建聚类树ꎬ以基因数量≥50 划分模块且 0.25 为模 值后逐渐下降ꎻ而 6 ̄BA 与 2ꎬ4 ̄D 协同处理组则呈
块合并阈值ꎬ最终得到 16 个基因模块( 图 6:B)ꎬ 现持续上升趋势ꎬ在第 21 天时活性最高ꎻ 当 3 种