
Page 8 - 《广西植物》2023年第9期
P. 8
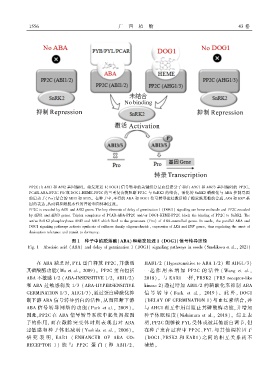
1 5 5 6 广 西 植 物 43 卷
PP2C 由 ABI1 和 ABI2 基因编码ꎮ 萌发延迟 1(DOG1)信号转导的关键组分是血红素分子和由 AHG1 和 AHG3 基因编码的 PP2Cꎮ
PCAR ̄ABA ̄PP2C 和 / 或 DOG1 ̄HEME ̄PP2C 的三重复合物阻断 PP2C 与 SnRK2 的结合ꎮ 活化的 SnRK2 磷酸化与 ABA 控制基因
的启动子(Pro)结合的 ABI3 和 ABI5ꎮ 在种子中ꎬ平行的 ABA 和 DOG1 信号转导途径激活棉子糖家族寡糖的合成、LEA 和 HSP 基
因的表达ꎬ从而调控耐脱水性的开始和向休眠过渡ꎮ
PP2C is encoded by ABI1 and ABI2 genes. The key elements of delay of germination 1 (DOG1) signaling are heme molecule and PP2C encoded
by AHG1 and AHG3 genes. Triplex complexes of PCAR ̄ABA ̄PP2C and/ or DOG1 ̄HEME ̄PP2C block the binding of PP2C to SnRK2. The
active SnRK2 phosphorylates ABI3 and ABI5 which bind to the promoters (Pro) of ABA ̄controlled genes. In seedsꎬ the parallel ABA and
DOG1 signaling pathways activate synthesis of raffinose family oligosaccharideꎬ expression of LEA and HSP genesꎬ thus regulating the onset of
desiccation tolerance and transit to dormancy.
图 1 种子中的脱落酸(ABA)和萌发延迟 1 (DOG1)信号转导途径
Fig. 1 Abscisic acid (ABA) and delay of germination 1 (DOG1) signaling pathways in seeds (Smolikova et al.ꎬ 2021)
在 ABA 缺乏时ꎬPYL 蛋白释放 PP2Cꎬ并激活 HAB1 / 2 (Hypersensitive to ABA 1 / 2) 和 AHG1 / 3)
其磷酸酶功能(Ma et al.ꎬ 2009)ꎮ PP2C 蛋白包括 一起 作 用 来 增 加 PP2C 的 活 性 ( Wang et al.ꎬ
ABA 不敏感 1 / 2 (ABA ̄INSENSITIVE 1 / 2ꎬ ABI1 / 2) 2018)ꎮ 与 EAR1 一样ꎬPR5K2 ( PR5 receptor ̄like
和 ABA 过敏感萌发 1 / 3 ( ABA ̄HYPERSENSITIVE kinase 2) 通过增加 ABI1 / 2 的磷酸化来抑制 ABA
GERMINATION 1 / 3ꎬ AHG1 / 3)ꎬ通过蛋白磷酸化抑 信 号 转 导 ( Baek et al.ꎬ 2019 )ꎮ 此 外ꎬ DOG1
制下游 ABA 信号转导蛋白的活性ꎬ从而阻断下游 (DELAY OF GERMINATION 1) 与血红素结合ꎬ并
ABA 信 号 转 导 网 络 的 功 能 ( Park et al.ꎬ 2009)ꎮ 与 AHG1 相互作用以阻止其磷酸酶功能ꎬ并增加
因此ꎬPP2C 在 ABA 信号转导系统中起负调控因 种子休眠程度( Nishimura et al.ꎬ 2018)ꎮ 综上表
子的作 用ꎬ 而 在 敲 除 突 变 体 时 则 表 现 出 对 ABA 明ꎬPP2C 能够被 PYL 受体或被其他蛋白调节ꎬ但
过敏感 和 种 子 休 眠 减 弱 ( Yoshida et al.ꎬ 2006)ꎮ 在种子发育过程中 PP2C、PYL 与其他调控因子
研 究 表 明ꎬ EAR1 ( ENHANCER OF ABA CO ̄ (DOG1、PR5K2 和 EAR1) 之 间 的 相 互 关 系 尚 不
RECEPTOR 1 ) 能 与 PP2C 蛋 白 ( 即 ABI1 / 2、 清楚ꎮ