
Page 10 - 《广西植物》2023年第9期
P. 10
1 5 5 8 广 西 植 物 43 卷
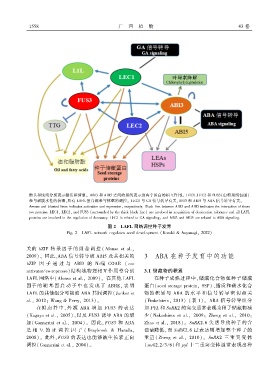
箭头和钝线分别表示激活和抑制ꎮ ABI3 和 ABI5 之间的黑线表示这两个蛋白的相互作用ꎮ LEC1、LEC2 和 FUS3(由粗黑线包围)
参与耐脱水性的获得ꎬ所有 LAFL 蛋白都参与休眠的调控ꎮ LEC1 与 GA 信号转导有关ꎬABI3 和 ABI5 与 ABA 信号转导有关ꎮ
Arrows and blunted lines indicates activation and repressionꎬ respectively. Black line between ABI3 and ABI5 indicates the interaction of these
two proteins. LEC1ꎬ LEC2ꎬ and FUS3 (surrounded by the thick black line) are involved in acquisition of desiccation tolerance and all LAFL
proteins are involved in the regulation of dormancy. LEC1 is related to GA signalingꎬ and ABI3 and ABI5 are related to ABA signaling.
图 2 LAFL 网络调控种子发育
Fig. 2 LAFL network regulates seed development (Kozaki & Aoyanagiꎬ 2022)
关的 bZIP 转 录 因 子 的 组 合 调 控 ( Alonso et al.ꎬ
2009)ꎮ 因此ꎬABA 信号转导被 ABI5 及其相关的 3 ABA 在种子发育中的功能
bZIP 因 子 通 过 与 ABI3 的 N ̄端 COAR ( co ̄
activator / co ̄represso)结构域物理相互作用整合到 3.1 储藏物的积累
LAFL 网络中( Alonso et al.ꎬ 2009)ꎮ 在其他 LAFL 在种子成熟过程中ꎬ储藏化合物如种子储藏
因子 的 靶 基 因 启 动 子 中 也 发 现 了 ABREꎬ 表 明 蛋白(seed storage proteinꎬ SSP)、脂质和碳水化合
LAFL 的其他组分可能被 ABA 共同调控( Junker et 物的积 累 与 ABA 的 水 平 和 信 号 转 导 密 切 相 关
al.ꎬ 2012ꎻ Wang & Perryꎬ 2013)ꎮ (Finkelsteinꎬ 2013) ( 表 1)ꎮ ABA 信号转导组分
在拟 南 芥 中ꎬ 外 源 ABA 增 加 FUS3 的 表 达 如 PYL 和 SnRK2 的突变通常表现为种子储藏物减
(Kagaya et al.ꎬ 2005)ꎬ以及 FUS3 诱导 ABA 的增 少( Nakashima et al.ꎬ 2009ꎻ Zheng et al.ꎬ 2010ꎻ
加(Gazzarrini et al.ꎬ 2004)ꎮ 因 此ꎬFUS3 和 ABA Zhao et al.ꎬ 2018)ꎮ SnRK2.6 失活导致种子的含
是 相 互 的 正 调 控 因 子 ( Braybrook & Haradaꎬ 油量降低ꎬ而 SnRK2.6 过表达则增加整个种子的
2008)ꎮ 此外ꎬFUS3 的表达也能够被生长素正向 重量 ( Zheng et al.ꎬ 2010)ꎮ SnRK2 三 重 突 变 体
调控(Gazzarrini et al.ꎬ 2004)ꎮ (snrk2.2 / 3 / 6) 和 pyl 十二重突变体通常表现出种