
Page 169 - 《广西植物》2024年第5期
P. 169
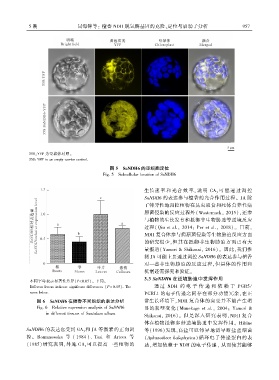
5 期 闫海锋等: 檀香 NDH 脱氢酶基因的克隆、定位与启动子分析 9 5 7
35S:YFP 为空载体对照ꎮ
35S:YFP is an empty carrier control.
图 5 SaNDH6 的亚细胞定位
Fig. 5 Subcellular location of SaNDH6
生长速 率 和 光 合 效 率ꎬ 说 明 GA 可 能 通 过 调 控
3
SaNDH6 的表达参与檀香的光合作用过程ꎮ JA 除
了特异性地调控植物在昆虫取食和死体营养性病
原菌侵染的反应过程外( Wasternackꎬ 2015)ꎬ还参
与植物的生长发育和抵御非生物胁迫等逆境反应
过程( Qiu et al.ꎬ 2014ꎻ Per et al.ꎬ 2018)ꎮ 目前ꎬ
NDH 复合体参与病原菌侵染等生物胁迫反应方面
的研究很少ꎬ但其在抵御非生物胁迫方面已有大
量报道(Yamori & Shikanaiꎬ 2016)ꎮ 因此ꎬ我们推
测 JA 可能主要通过调控 SaNDH6 的表达参与檀香
对一些非生物胁迫的反应过程ꎬ但具体的作用和
机制还需探究和验证ꎮ
3.3 SaNDH6 在逆境胁迫中发挥作用
不同字母表示显著性差异(P<0.05)ꎮ 下同ꎮ
Different letters indicate significant differences (P < 0.05). The 通 过 NDH 的 电 子 传 递 和 依 赖 于 PGR5 /
same below. PGRL1 的电子传递之间存在部分功能冗余ꎬ在正
图 6 SaNDH6 在檀香不同组织的表达分析 常生长环境下ꎬNDH 复合体的突变并不能产生明
Fig. 6 Relative expression analysis of SaDNH6 显的表型变化( Munekage et al.ꎬ 2004ꎻ Yamori &
in different tissues of Santalum album
Shikanaiꎬ 2016)ꎮ 但是深入研究表明ꎬNDH 复合
体在植物抵御多种逆境胁迫中发挥作用ꎮ Hibino
SaNDH6 的表达也受到 GA 和 JA 等激素的正向调 等(1996)发现ꎬ高盐可以特异地诱导耐盐蓝细菌
3
控ꎮ Romanowska 等 ( 1984 )、 Tsai 和 Arteca 等 (Aphanothece halophytica) 循环电子传递蛋白的表
(1985)研究表明ꎬ外施 GA 可以提高一些植物的 达ꎬ增加依赖于 NDH 的电子传递ꎬ 从而使其能够
3